User login
CCJM delivers practical clinical articles relevant to internists, cardiologists, endocrinologists, and other specialists, all written by known experts.
Copyright © 2019 Cleveland Clinic. All rights reserved. The information provided is for educational purposes only. Use of this website is subject to the disclaimer and privacy policy.
gambling
compulsive behaviors
ammunition
assault rifle
black jack
Boko Haram
bondage
child abuse
cocaine
Daech
drug paraphernalia
explosion
gun
human trafficking
ISIL
ISIS
Islamic caliphate
Islamic state
mixed martial arts
MMA
molestation
national rifle association
NRA
nsfw
pedophile
pedophilia
poker
porn
pornography
psychedelic drug
recreational drug
sex slave rings
slot machine
terrorism
terrorist
Texas hold 'em
UFC
substance abuse
abuseed
abuseer
abusees
abuseing
abusely
abuses
aeolus
aeolused
aeoluser
aeoluses
aeolusing
aeolusly
aeoluss
ahole
aholeed
aholeer
aholees
aholeing
aholely
aholes
alcohol
alcoholed
alcoholer
alcoholes
alcoholing
alcoholly
alcohols
allman
allmaned
allmaner
allmanes
allmaning
allmanly
allmans
alted
altes
alting
altly
alts
analed
analer
anales
analing
anally
analprobe
analprobeed
analprobeer
analprobees
analprobeing
analprobely
analprobes
anals
anilingus
anilingused
anilinguser
anilinguses
anilingusing
anilingusly
anilinguss
anus
anused
anuser
anuses
anusing
anusly
anuss
areola
areolaed
areolaer
areolaes
areolaing
areolaly
areolas
areole
areoleed
areoleer
areolees
areoleing
areolely
areoles
arian
arianed
arianer
arianes
arianing
arianly
arians
aryan
aryaned
aryaner
aryanes
aryaning
aryanly
aryans
asiaed
asiaer
asiaes
asiaing
asialy
asias
ass
ass hole
ass lick
ass licked
ass licker
ass lickes
ass licking
ass lickly
ass licks
assbang
assbanged
assbangeded
assbangeder
assbangedes
assbangeding
assbangedly
assbangeds
assbanger
assbanges
assbanging
assbangly
assbangs
assbangsed
assbangser
assbangses
assbangsing
assbangsly
assbangss
assed
asser
asses
assesed
asseser
asseses
assesing
assesly
assess
assfuck
assfucked
assfucker
assfuckered
assfuckerer
assfuckeres
assfuckering
assfuckerly
assfuckers
assfuckes
assfucking
assfuckly
assfucks
asshat
asshated
asshater
asshates
asshating
asshatly
asshats
assholeed
assholeer
assholees
assholeing
assholely
assholes
assholesed
assholeser
assholeses
assholesing
assholesly
assholess
assing
assly
assmaster
assmastered
assmasterer
assmasteres
assmastering
assmasterly
assmasters
assmunch
assmunched
assmuncher
assmunches
assmunching
assmunchly
assmunchs
asss
asswipe
asswipeed
asswipeer
asswipees
asswipeing
asswipely
asswipes
asswipesed
asswipeser
asswipeses
asswipesing
asswipesly
asswipess
azz
azzed
azzer
azzes
azzing
azzly
azzs
babeed
babeer
babees
babeing
babely
babes
babesed
babeser
babeses
babesing
babesly
babess
ballsac
ballsaced
ballsacer
ballsaces
ballsacing
ballsack
ballsacked
ballsacker
ballsackes
ballsacking
ballsackly
ballsacks
ballsacly
ballsacs
ballsed
ballser
ballses
ballsing
ballsly
ballss
barf
barfed
barfer
barfes
barfing
barfly
barfs
bastard
bastarded
bastarder
bastardes
bastarding
bastardly
bastards
bastardsed
bastardser
bastardses
bastardsing
bastardsly
bastardss
bawdy
bawdyed
bawdyer
bawdyes
bawdying
bawdyly
bawdys
beaner
beanered
beanerer
beaneres
beanering
beanerly
beaners
beardedclam
beardedclamed
beardedclamer
beardedclames
beardedclaming
beardedclamly
beardedclams
beastiality
beastialityed
beastialityer
beastialityes
beastialitying
beastialityly
beastialitys
beatch
beatched
beatcher
beatches
beatching
beatchly
beatchs
beater
beatered
beaterer
beateres
beatering
beaterly
beaters
beered
beerer
beeres
beering
beerly
beeyotch
beeyotched
beeyotcher
beeyotches
beeyotching
beeyotchly
beeyotchs
beotch
beotched
beotcher
beotches
beotching
beotchly
beotchs
biatch
biatched
biatcher
biatches
biatching
biatchly
biatchs
big tits
big titsed
big titser
big titses
big titsing
big titsly
big titss
bigtits
bigtitsed
bigtitser
bigtitses
bigtitsing
bigtitsly
bigtitss
bimbo
bimboed
bimboer
bimboes
bimboing
bimboly
bimbos
bisexualed
bisexualer
bisexuales
bisexualing
bisexually
bisexuals
bitch
bitched
bitcheded
bitcheder
bitchedes
bitcheding
bitchedly
bitcheds
bitcher
bitches
bitchesed
bitcheser
bitcheses
bitchesing
bitchesly
bitchess
bitching
bitchly
bitchs
bitchy
bitchyed
bitchyer
bitchyes
bitchying
bitchyly
bitchys
bleached
bleacher
bleaches
bleaching
bleachly
bleachs
blow job
blow jobed
blow jober
blow jobes
blow jobing
blow jobly
blow jobs
blowed
blower
blowes
blowing
blowjob
blowjobed
blowjober
blowjobes
blowjobing
blowjobly
blowjobs
blowjobsed
blowjobser
blowjobses
blowjobsing
blowjobsly
blowjobss
blowly
blows
boink
boinked
boinker
boinkes
boinking
boinkly
boinks
bollock
bollocked
bollocker
bollockes
bollocking
bollockly
bollocks
bollocksed
bollockser
bollockses
bollocksing
bollocksly
bollockss
bollok
bolloked
bolloker
bollokes
bolloking
bollokly
bolloks
boner
bonered
bonerer
boneres
bonering
bonerly
boners
bonersed
bonerser
bonerses
bonersing
bonersly
bonerss
bong
bonged
bonger
bonges
bonging
bongly
bongs
boob
boobed
boober
boobes
boobies
boobiesed
boobieser
boobieses
boobiesing
boobiesly
boobiess
boobing
boobly
boobs
boobsed
boobser
boobses
boobsing
boobsly
boobss
booby
boobyed
boobyer
boobyes
boobying
boobyly
boobys
booger
boogered
boogerer
boogeres
boogering
boogerly
boogers
bookie
bookieed
bookieer
bookiees
bookieing
bookiely
bookies
bootee
booteeed
booteeer
booteees
booteeing
booteely
bootees
bootie
bootieed
bootieer
bootiees
bootieing
bootiely
booties
booty
bootyed
bootyer
bootyes
bootying
bootyly
bootys
boozeed
boozeer
boozees
boozeing
boozely
boozer
boozered
boozerer
boozeres
boozering
boozerly
boozers
boozes
boozy
boozyed
boozyer
boozyes
boozying
boozyly
boozys
bosomed
bosomer
bosomes
bosoming
bosomly
bosoms
bosomy
bosomyed
bosomyer
bosomyes
bosomying
bosomyly
bosomys
bugger
buggered
buggerer
buggeres
buggering
buggerly
buggers
bukkake
bukkakeed
bukkakeer
bukkakees
bukkakeing
bukkakely
bukkakes
bull shit
bull shited
bull shiter
bull shites
bull shiting
bull shitly
bull shits
bullshit
bullshited
bullshiter
bullshites
bullshiting
bullshitly
bullshits
bullshitsed
bullshitser
bullshitses
bullshitsing
bullshitsly
bullshitss
bullshitted
bullshitteded
bullshitteder
bullshittedes
bullshitteding
bullshittedly
bullshitteds
bullturds
bullturdsed
bullturdser
bullturdses
bullturdsing
bullturdsly
bullturdss
bung
bunged
bunger
bunges
bunging
bungly
bungs
busty
bustyed
bustyer
bustyes
bustying
bustyly
bustys
butt
butt fuck
butt fucked
butt fucker
butt fuckes
butt fucking
butt fuckly
butt fucks
butted
buttes
buttfuck
buttfucked
buttfucker
buttfuckered
buttfuckerer
buttfuckeres
buttfuckering
buttfuckerly
buttfuckers
buttfuckes
buttfucking
buttfuckly
buttfucks
butting
buttly
buttplug
buttpluged
buttpluger
buttpluges
buttpluging
buttplugly
buttplugs
butts
caca
cacaed
cacaer
cacaes
cacaing
cacaly
cacas
cahone
cahoneed
cahoneer
cahonees
cahoneing
cahonely
cahones
cameltoe
cameltoeed
cameltoeer
cameltoees
cameltoeing
cameltoely
cameltoes
carpetmuncher
carpetmunchered
carpetmuncherer
carpetmuncheres
carpetmunchering
carpetmuncherly
carpetmunchers
cawk
cawked
cawker
cawkes
cawking
cawkly
cawks
chinc
chinced
chincer
chinces
chincing
chincly
chincs
chincsed
chincser
chincses
chincsing
chincsly
chincss
chink
chinked
chinker
chinkes
chinking
chinkly
chinks
chode
chodeed
chodeer
chodees
chodeing
chodely
chodes
chodesed
chodeser
chodeses
chodesing
chodesly
chodess
clit
clited
cliter
clites
cliting
clitly
clitoris
clitorised
clitoriser
clitorises
clitorising
clitorisly
clitoriss
clitorus
clitorused
clitoruser
clitoruses
clitorusing
clitorusly
clitoruss
clits
clitsed
clitser
clitses
clitsing
clitsly
clitss
clitty
clittyed
clittyer
clittyes
clittying
clittyly
clittys
cocain
cocaine
cocained
cocaineed
cocaineer
cocainees
cocaineing
cocainely
cocainer
cocaines
cocaining
cocainly
cocains
cock
cock sucker
cock suckered
cock suckerer
cock suckeres
cock suckering
cock suckerly
cock suckers
cockblock
cockblocked
cockblocker
cockblockes
cockblocking
cockblockly
cockblocks
cocked
cocker
cockes
cockholster
cockholstered
cockholsterer
cockholsteres
cockholstering
cockholsterly
cockholsters
cocking
cockknocker
cockknockered
cockknockerer
cockknockeres
cockknockering
cockknockerly
cockknockers
cockly
cocks
cocksed
cockser
cockses
cocksing
cocksly
cocksmoker
cocksmokered
cocksmokerer
cocksmokeres
cocksmokering
cocksmokerly
cocksmokers
cockss
cocksucker
cocksuckered
cocksuckerer
cocksuckeres
cocksuckering
cocksuckerly
cocksuckers
coital
coitaled
coitaler
coitales
coitaling
coitally
coitals
commie
commieed
commieer
commiees
commieing
commiely
commies
condomed
condomer
condomes
condoming
condomly
condoms
coon
cooned
cooner
coones
cooning
coonly
coons
coonsed
coonser
coonses
coonsing
coonsly
coonss
corksucker
corksuckered
corksuckerer
corksuckeres
corksuckering
corksuckerly
corksuckers
cracked
crackwhore
crackwhoreed
crackwhoreer
crackwhorees
crackwhoreing
crackwhorely
crackwhores
crap
craped
craper
crapes
craping
craply
crappy
crappyed
crappyer
crappyes
crappying
crappyly
crappys
cum
cumed
cumer
cumes
cuming
cumly
cummin
cummined
cumminer
cummines
cumming
cumminged
cumminger
cumminges
cumminging
cummingly
cummings
cummining
cumminly
cummins
cums
cumshot
cumshoted
cumshoter
cumshotes
cumshoting
cumshotly
cumshots
cumshotsed
cumshotser
cumshotses
cumshotsing
cumshotsly
cumshotss
cumslut
cumsluted
cumsluter
cumslutes
cumsluting
cumslutly
cumsluts
cumstain
cumstained
cumstainer
cumstaines
cumstaining
cumstainly
cumstains
cunilingus
cunilingused
cunilinguser
cunilinguses
cunilingusing
cunilingusly
cunilinguss
cunnilingus
cunnilingused
cunnilinguser
cunnilinguses
cunnilingusing
cunnilingusly
cunnilinguss
cunny
cunnyed
cunnyer
cunnyes
cunnying
cunnyly
cunnys
cunt
cunted
cunter
cuntes
cuntface
cuntfaceed
cuntfaceer
cuntfacees
cuntfaceing
cuntfacely
cuntfaces
cunthunter
cunthuntered
cunthunterer
cunthunteres
cunthuntering
cunthunterly
cunthunters
cunting
cuntlick
cuntlicked
cuntlicker
cuntlickered
cuntlickerer
cuntlickeres
cuntlickering
cuntlickerly
cuntlickers
cuntlickes
cuntlicking
cuntlickly
cuntlicks
cuntly
cunts
cuntsed
cuntser
cuntses
cuntsing
cuntsly
cuntss
dago
dagoed
dagoer
dagoes
dagoing
dagoly
dagos
dagosed
dagoser
dagoses
dagosing
dagosly
dagoss
dammit
dammited
dammiter
dammites
dammiting
dammitly
dammits
damn
damned
damneded
damneder
damnedes
damneding
damnedly
damneds
damner
damnes
damning
damnit
damnited
damniter
damnites
damniting
damnitly
damnits
damnly
damns
dick
dickbag
dickbaged
dickbager
dickbages
dickbaging
dickbagly
dickbags
dickdipper
dickdippered
dickdipperer
dickdipperes
dickdippering
dickdipperly
dickdippers
dicked
dicker
dickes
dickface
dickfaceed
dickfaceer
dickfacees
dickfaceing
dickfacely
dickfaces
dickflipper
dickflippered
dickflipperer
dickflipperes
dickflippering
dickflipperly
dickflippers
dickhead
dickheaded
dickheader
dickheades
dickheading
dickheadly
dickheads
dickheadsed
dickheadser
dickheadses
dickheadsing
dickheadsly
dickheadss
dicking
dickish
dickished
dickisher
dickishes
dickishing
dickishly
dickishs
dickly
dickripper
dickrippered
dickripperer
dickripperes
dickrippering
dickripperly
dickrippers
dicks
dicksipper
dicksippered
dicksipperer
dicksipperes
dicksippering
dicksipperly
dicksippers
dickweed
dickweeded
dickweeder
dickweedes
dickweeding
dickweedly
dickweeds
dickwhipper
dickwhippered
dickwhipperer
dickwhipperes
dickwhippering
dickwhipperly
dickwhippers
dickzipper
dickzippered
dickzipperer
dickzipperes
dickzippering
dickzipperly
dickzippers
diddle
diddleed
diddleer
diddlees
diddleing
diddlely
diddles
dike
dikeed
dikeer
dikees
dikeing
dikely
dikes
dildo
dildoed
dildoer
dildoes
dildoing
dildoly
dildos
dildosed
dildoser
dildoses
dildosing
dildosly
dildoss
diligaf
diligafed
diligafer
diligafes
diligafing
diligafly
diligafs
dillweed
dillweeded
dillweeder
dillweedes
dillweeding
dillweedly
dillweeds
dimwit
dimwited
dimwiter
dimwites
dimwiting
dimwitly
dimwits
dingle
dingleed
dingleer
dinglees
dingleing
dinglely
dingles
dipship
dipshiped
dipshiper
dipshipes
dipshiping
dipshiply
dipships
dizzyed
dizzyer
dizzyes
dizzying
dizzyly
dizzys
doggiestyleed
doggiestyleer
doggiestylees
doggiestyleing
doggiestylely
doggiestyles
doggystyleed
doggystyleer
doggystylees
doggystyleing
doggystylely
doggystyles
dong
donged
donger
donges
donging
dongly
dongs
doofus
doofused
doofuser
doofuses
doofusing
doofusly
doofuss
doosh
dooshed
doosher
dooshes
dooshing
dooshly
dooshs
dopeyed
dopeyer
dopeyes
dopeying
dopeyly
dopeys
douchebag
douchebaged
douchebager
douchebages
douchebaging
douchebagly
douchebags
douchebagsed
douchebagser
douchebagses
douchebagsing
douchebagsly
douchebagss
doucheed
doucheer
douchees
doucheing
douchely
douches
douchey
doucheyed
doucheyer
doucheyes
doucheying
doucheyly
doucheys
drunk
drunked
drunker
drunkes
drunking
drunkly
drunks
dumass
dumassed
dumasser
dumasses
dumassing
dumassly
dumasss
dumbass
dumbassed
dumbasser
dumbasses
dumbassesed
dumbasseser
dumbasseses
dumbassesing
dumbassesly
dumbassess
dumbassing
dumbassly
dumbasss
dummy
dummyed
dummyer
dummyes
dummying
dummyly
dummys
dyke
dykeed
dykeer
dykees
dykeing
dykely
dykes
dykesed
dykeser
dykeses
dykesing
dykesly
dykess
erotic
eroticed
eroticer
erotices
eroticing
eroticly
erotics
extacy
extacyed
extacyer
extacyes
extacying
extacyly
extacys
extasy
extasyed
extasyer
extasyes
extasying
extasyly
extasys
fack
facked
facker
fackes
facking
fackly
facks
fag
faged
fager
fages
fagg
fagged
faggeded
faggeder
faggedes
faggeding
faggedly
faggeds
fagger
fagges
fagging
faggit
faggited
faggiter
faggites
faggiting
faggitly
faggits
faggly
faggot
faggoted
faggoter
faggotes
faggoting
faggotly
faggots
faggs
faging
fagly
fagot
fagoted
fagoter
fagotes
fagoting
fagotly
fagots
fags
fagsed
fagser
fagses
fagsing
fagsly
fagss
faig
faiged
faiger
faiges
faiging
faigly
faigs
faigt
faigted
faigter
faigtes
faigting
faigtly
faigts
fannybandit
fannybandited
fannybanditer
fannybandites
fannybanditing
fannybanditly
fannybandits
farted
farter
fartes
farting
fartknocker
fartknockered
fartknockerer
fartknockeres
fartknockering
fartknockerly
fartknockers
fartly
farts
felch
felched
felcher
felchered
felcherer
felcheres
felchering
felcherly
felchers
felches
felching
felchinged
felchinger
felchinges
felchinging
felchingly
felchings
felchly
felchs
fellate
fellateed
fellateer
fellatees
fellateing
fellately
fellates
fellatio
fellatioed
fellatioer
fellatioes
fellatioing
fellatioly
fellatios
feltch
feltched
feltcher
feltchered
feltcherer
feltcheres
feltchering
feltcherly
feltchers
feltches
feltching
feltchly
feltchs
feom
feomed
feomer
feomes
feoming
feomly
feoms
fisted
fisteded
fisteder
fistedes
fisteding
fistedly
fisteds
fisting
fistinged
fistinger
fistinges
fistinging
fistingly
fistings
fisty
fistyed
fistyer
fistyes
fistying
fistyly
fistys
floozy
floozyed
floozyer
floozyes
floozying
floozyly
floozys
foad
foaded
foader
foades
foading
foadly
foads
fondleed
fondleer
fondlees
fondleing
fondlely
fondles
foobar
foobared
foobarer
foobares
foobaring
foobarly
foobars
freex
freexed
freexer
freexes
freexing
freexly
freexs
frigg
frigga
friggaed
friggaer
friggaes
friggaing
friggaly
friggas
frigged
frigger
frigges
frigging
friggly
friggs
fubar
fubared
fubarer
fubares
fubaring
fubarly
fubars
fuck
fuckass
fuckassed
fuckasser
fuckasses
fuckassing
fuckassly
fuckasss
fucked
fuckeded
fuckeder
fuckedes
fuckeding
fuckedly
fuckeds
fucker
fuckered
fuckerer
fuckeres
fuckering
fuckerly
fuckers
fuckes
fuckface
fuckfaceed
fuckfaceer
fuckfacees
fuckfaceing
fuckfacely
fuckfaces
fuckin
fuckined
fuckiner
fuckines
fucking
fuckinged
fuckinger
fuckinges
fuckinging
fuckingly
fuckings
fuckining
fuckinly
fuckins
fuckly
fucknugget
fucknuggeted
fucknuggeter
fucknuggetes
fucknuggeting
fucknuggetly
fucknuggets
fucknut
fucknuted
fucknuter
fucknutes
fucknuting
fucknutly
fucknuts
fuckoff
fuckoffed
fuckoffer
fuckoffes
fuckoffing
fuckoffly
fuckoffs
fucks
fucksed
fuckser
fuckses
fucksing
fucksly
fuckss
fucktard
fucktarded
fucktarder
fucktardes
fucktarding
fucktardly
fucktards
fuckup
fuckuped
fuckuper
fuckupes
fuckuping
fuckuply
fuckups
fuckwad
fuckwaded
fuckwader
fuckwades
fuckwading
fuckwadly
fuckwads
fuckwit
fuckwited
fuckwiter
fuckwites
fuckwiting
fuckwitly
fuckwits
fudgepacker
fudgepackered
fudgepackerer
fudgepackeres
fudgepackering
fudgepackerly
fudgepackers
fuk
fuked
fuker
fukes
fuking
fukly
fuks
fvck
fvcked
fvcker
fvckes
fvcking
fvckly
fvcks
fxck
fxcked
fxcker
fxckes
fxcking
fxckly
fxcks
gae
gaeed
gaeer
gaees
gaeing
gaely
gaes
gai
gaied
gaier
gaies
gaiing
gaily
gais
ganja
ganjaed
ganjaer
ganjaes
ganjaing
ganjaly
ganjas
gayed
gayer
gayes
gaying
gayly
gays
gaysed
gayser
gayses
gaysing
gaysly
gayss
gey
geyed
geyer
geyes
geying
geyly
geys
gfc
gfced
gfcer
gfces
gfcing
gfcly
gfcs
gfy
gfyed
gfyer
gfyes
gfying
gfyly
gfys
ghay
ghayed
ghayer
ghayes
ghaying
ghayly
ghays
ghey
gheyed
gheyer
gheyes
gheying
gheyly
gheys
gigolo
gigoloed
gigoloer
gigoloes
gigoloing
gigololy
gigolos
goatse
goatseed
goatseer
goatsees
goatseing
goatsely
goatses
godamn
godamned
godamner
godamnes
godamning
godamnit
godamnited
godamniter
godamnites
godamniting
godamnitly
godamnits
godamnly
godamns
goddam
goddamed
goddamer
goddames
goddaming
goddamly
goddammit
goddammited
goddammiter
goddammites
goddammiting
goddammitly
goddammits
goddamn
goddamned
goddamner
goddamnes
goddamning
goddamnly
goddamns
goddams
goldenshower
goldenshowered
goldenshowerer
goldenshoweres
goldenshowering
goldenshowerly
goldenshowers
gonad
gonaded
gonader
gonades
gonading
gonadly
gonads
gonadsed
gonadser
gonadses
gonadsing
gonadsly
gonadss
gook
gooked
gooker
gookes
gooking
gookly
gooks
gooksed
gookser
gookses
gooksing
gooksly
gookss
gringo
gringoed
gringoer
gringoes
gringoing
gringoly
gringos
gspot
gspoted
gspoter
gspotes
gspoting
gspotly
gspots
gtfo
gtfoed
gtfoer
gtfoes
gtfoing
gtfoly
gtfos
guido
guidoed
guidoer
guidoes
guidoing
guidoly
guidos
handjob
handjobed
handjober
handjobes
handjobing
handjobly
handjobs
hard on
hard oned
hard oner
hard ones
hard oning
hard only
hard ons
hardknight
hardknighted
hardknighter
hardknightes
hardknighting
hardknightly
hardknights
hebe
hebeed
hebeer
hebees
hebeing
hebely
hebes
heeb
heebed
heeber
heebes
heebing
heebly
heebs
hell
helled
heller
helles
helling
hellly
hells
hemp
hemped
hemper
hempes
hemping
hemply
hemps
heroined
heroiner
heroines
heroining
heroinly
heroins
herp
herped
herper
herpes
herpesed
herpeser
herpeses
herpesing
herpesly
herpess
herping
herply
herps
herpy
herpyed
herpyer
herpyes
herpying
herpyly
herpys
hitler
hitlered
hitlerer
hitleres
hitlering
hitlerly
hitlers
hived
hiver
hives
hiving
hivly
hivs
hobag
hobaged
hobager
hobages
hobaging
hobagly
hobags
homey
homeyed
homeyer
homeyes
homeying
homeyly
homeys
homo
homoed
homoer
homoes
homoey
homoeyed
homoeyer
homoeyes
homoeying
homoeyly
homoeys
homoing
homoly
homos
honky
honkyed
honkyer
honkyes
honkying
honkyly
honkys
hooch
hooched
hoocher
hooches
hooching
hoochly
hoochs
hookah
hookahed
hookaher
hookahes
hookahing
hookahly
hookahs
hooker
hookered
hookerer
hookeres
hookering
hookerly
hookers
hoor
hoored
hoorer
hoores
hooring
hoorly
hoors
hootch
hootched
hootcher
hootches
hootching
hootchly
hootchs
hooter
hootered
hooterer
hooteres
hootering
hooterly
hooters
hootersed
hooterser
hooterses
hootersing
hootersly
hooterss
horny
hornyed
hornyer
hornyes
hornying
hornyly
hornys
houstoned
houstoner
houstones
houstoning
houstonly
houstons
hump
humped
humpeded
humpeder
humpedes
humpeding
humpedly
humpeds
humper
humpes
humping
humpinged
humpinger
humpinges
humpinging
humpingly
humpings
humply
humps
husbanded
husbander
husbandes
husbanding
husbandly
husbands
hussy
hussyed
hussyer
hussyes
hussying
hussyly
hussys
hymened
hymener
hymenes
hymening
hymenly
hymens
inbred
inbreded
inbreder
inbredes
inbreding
inbredly
inbreds
incest
incested
incester
incestes
incesting
incestly
incests
injun
injuned
injuner
injunes
injuning
injunly
injuns
jackass
jackassed
jackasser
jackasses
jackassing
jackassly
jackasss
jackhole
jackholeed
jackholeer
jackholees
jackholeing
jackholely
jackholes
jackoff
jackoffed
jackoffer
jackoffes
jackoffing
jackoffly
jackoffs
jap
japed
japer
japes
japing
japly
japs
japsed
japser
japses
japsing
japsly
japss
jerkoff
jerkoffed
jerkoffer
jerkoffes
jerkoffing
jerkoffly
jerkoffs
jerks
jism
jismed
jismer
jismes
jisming
jismly
jisms
jiz
jized
jizer
jizes
jizing
jizly
jizm
jizmed
jizmer
jizmes
jizming
jizmly
jizms
jizs
jizz
jizzed
jizzeded
jizzeder
jizzedes
jizzeding
jizzedly
jizzeds
jizzer
jizzes
jizzing
jizzly
jizzs
junkie
junkieed
junkieer
junkiees
junkieing
junkiely
junkies
junky
junkyed
junkyer
junkyes
junkying
junkyly
junkys
kike
kikeed
kikeer
kikees
kikeing
kikely
kikes
kikesed
kikeser
kikeses
kikesing
kikesly
kikess
killed
killer
killes
killing
killly
kills
kinky
kinkyed
kinkyer
kinkyes
kinkying
kinkyly
kinkys
kkk
kkked
kkker
kkkes
kkking
kkkly
kkks
klan
klaned
klaner
klanes
klaning
klanly
klans
knobend
knobended
knobender
knobendes
knobending
knobendly
knobends
kooch
kooched
koocher
kooches
koochesed
koocheser
koocheses
koochesing
koochesly
koochess
kooching
koochly
koochs
kootch
kootched
kootcher
kootches
kootching
kootchly
kootchs
kraut
krauted
krauter
krautes
krauting
krautly
krauts
kyke
kykeed
kykeer
kykees
kykeing
kykely
kykes
lech
leched
lecher
leches
leching
lechly
lechs
leper
lepered
leperer
leperes
lepering
leperly
lepers
lesbiansed
lesbianser
lesbianses
lesbiansing
lesbiansly
lesbianss
lesbo
lesboed
lesboer
lesboes
lesboing
lesboly
lesbos
lesbosed
lesboser
lesboses
lesbosing
lesbosly
lesboss
lez
lezbianed
lezbianer
lezbianes
lezbianing
lezbianly
lezbians
lezbiansed
lezbianser
lezbianses
lezbiansing
lezbiansly
lezbianss
lezbo
lezboed
lezboer
lezboes
lezboing
lezboly
lezbos
lezbosed
lezboser
lezboses
lezbosing
lezbosly
lezboss
lezed
lezer
lezes
lezing
lezly
lezs
lezzie
lezzieed
lezzieer
lezziees
lezzieing
lezziely
lezzies
lezziesed
lezzieser
lezzieses
lezziesing
lezziesly
lezziess
lezzy
lezzyed
lezzyer
lezzyes
lezzying
lezzyly
lezzys
lmaoed
lmaoer
lmaoes
lmaoing
lmaoly
lmaos
lmfao
lmfaoed
lmfaoer
lmfaoes
lmfaoing
lmfaoly
lmfaos
loined
loiner
loines
loining
loinly
loins
loinsed
loinser
loinses
loinsing
loinsly
loinss
lubeed
lubeer
lubees
lubeing
lubely
lubes
lusty
lustyed
lustyer
lustyes
lustying
lustyly
lustys
massa
massaed
massaer
massaes
massaing
massaly
massas
masterbate
masterbateed
masterbateer
masterbatees
masterbateing
masterbately
masterbates
masterbating
masterbatinged
masterbatinger
masterbatinges
masterbatinging
masterbatingly
masterbatings
masterbation
masterbationed
masterbationer
masterbationes
masterbationing
masterbationly
masterbations
masturbate
masturbateed
masturbateer
masturbatees
masturbateing
masturbately
masturbates
masturbating
masturbatinged
masturbatinger
masturbatinges
masturbatinging
masturbatingly
masturbatings
masturbation
masturbationed
masturbationer
masturbationes
masturbationing
masturbationly
masturbations
methed
mether
methes
mething
methly
meths
militaryed
militaryer
militaryes
militarying
militaryly
militarys
mofo
mofoed
mofoer
mofoes
mofoing
mofoly
mofos
molest
molested
molester
molestes
molesting
molestly
molests
moolie
moolieed
moolieer
mooliees
moolieing
mooliely
moolies
moron
moroned
moroner
morones
moroning
moronly
morons
motherfucka
motherfuckaed
motherfuckaer
motherfuckaes
motherfuckaing
motherfuckaly
motherfuckas
motherfucker
motherfuckered
motherfuckerer
motherfuckeres
motherfuckering
motherfuckerly
motherfuckers
motherfucking
motherfuckinged
motherfuckinger
motherfuckinges
motherfuckinging
motherfuckingly
motherfuckings
mtherfucker
mtherfuckered
mtherfuckerer
mtherfuckeres
mtherfuckering
mtherfuckerly
mtherfuckers
mthrfucker
mthrfuckered
mthrfuckerer
mthrfuckeres
mthrfuckering
mthrfuckerly
mthrfuckers
mthrfucking
mthrfuckinged
mthrfuckinger
mthrfuckinges
mthrfuckinging
mthrfuckingly
mthrfuckings
muff
muffdiver
muffdivered
muffdiverer
muffdiveres
muffdivering
muffdiverly
muffdivers
muffed
muffer
muffes
muffing
muffly
muffs
murdered
murderer
murderes
murdering
murderly
murders
muthafuckaz
muthafuckazed
muthafuckazer
muthafuckazes
muthafuckazing
muthafuckazly
muthafuckazs
muthafucker
muthafuckered
muthafuckerer
muthafuckeres
muthafuckering
muthafuckerly
muthafuckers
mutherfucker
mutherfuckered
mutherfuckerer
mutherfuckeres
mutherfuckering
mutherfuckerly
mutherfuckers
mutherfucking
mutherfuckinged
mutherfuckinger
mutherfuckinges
mutherfuckinging
mutherfuckingly
mutherfuckings
muthrfucking
muthrfuckinged
muthrfuckinger
muthrfuckinges
muthrfuckinging
muthrfuckingly
muthrfuckings
nad
naded
nader
nades
nading
nadly
nads
nadsed
nadser
nadses
nadsing
nadsly
nadss
nakeded
nakeder
nakedes
nakeding
nakedly
nakeds
napalm
napalmed
napalmer
napalmes
napalming
napalmly
napalms
nappy
nappyed
nappyer
nappyes
nappying
nappyly
nappys
nazi
nazied
nazier
nazies
naziing
nazily
nazis
nazism
nazismed
nazismer
nazismes
nazisming
nazismly
nazisms
negro
negroed
negroer
negroes
negroing
negroly
negros
nigga
niggaed
niggaer
niggaes
niggah
niggahed
niggaher
niggahes
niggahing
niggahly
niggahs
niggaing
niggaly
niggas
niggased
niggaser
niggases
niggasing
niggasly
niggass
niggaz
niggazed
niggazer
niggazes
niggazing
niggazly
niggazs
nigger
niggered
niggerer
niggeres
niggering
niggerly
niggers
niggersed
niggerser
niggerses
niggersing
niggersly
niggerss
niggle
niggleed
niggleer
nigglees
niggleing
nigglely
niggles
niglet
nigleted
nigleter
nigletes
nigleting
nigletly
niglets
nimrod
nimroded
nimroder
nimrodes
nimroding
nimrodly
nimrods
ninny
ninnyed
ninnyer
ninnyes
ninnying
ninnyly
ninnys
nooky
nookyed
nookyer
nookyes
nookying
nookyly
nookys
nuccitelli
nuccitellied
nuccitellier
nuccitellies
nuccitelliing
nuccitellily
nuccitellis
nympho
nymphoed
nymphoer
nymphoes
nymphoing
nympholy
nymphos
opium
opiumed
opiumer
opiumes
opiuming
opiumly
opiums
orgies
orgiesed
orgieser
orgieses
orgiesing
orgiesly
orgiess
orgy
orgyed
orgyer
orgyes
orgying
orgyly
orgys
paddy
paddyed
paddyer
paddyes
paddying
paddyly
paddys
paki
pakied
pakier
pakies
pakiing
pakily
pakis
pantie
pantieed
pantieer
pantiees
pantieing
pantiely
panties
pantiesed
pantieser
pantieses
pantiesing
pantiesly
pantiess
panty
pantyed
pantyer
pantyes
pantying
pantyly
pantys
pastie
pastieed
pastieer
pastiees
pastieing
pastiely
pasties
pasty
pastyed
pastyer
pastyes
pastying
pastyly
pastys
pecker
peckered
peckerer
peckeres
peckering
peckerly
peckers
pedo
pedoed
pedoer
pedoes
pedoing
pedoly
pedophile
pedophileed
pedophileer
pedophilees
pedophileing
pedophilely
pedophiles
pedophilia
pedophiliac
pedophiliaced
pedophiliacer
pedophiliaces
pedophiliacing
pedophiliacly
pedophiliacs
pedophiliaed
pedophiliaer
pedophiliaes
pedophiliaing
pedophilialy
pedophilias
pedos
penial
penialed
penialer
peniales
penialing
penially
penials
penile
penileed
penileer
penilees
penileing
penilely
peniles
penis
penised
peniser
penises
penising
penisly
peniss
perversion
perversioned
perversioner
perversiones
perversioning
perversionly
perversions
peyote
peyoteed
peyoteer
peyotees
peyoteing
peyotely
peyotes
phuck
phucked
phucker
phuckes
phucking
phuckly
phucks
pillowbiter
pillowbitered
pillowbiterer
pillowbiteres
pillowbitering
pillowbiterly
pillowbiters
pimp
pimped
pimper
pimpes
pimping
pimply
pimps
pinko
pinkoed
pinkoer
pinkoes
pinkoing
pinkoly
pinkos
pissed
pisseded
pisseder
pissedes
pisseding
pissedly
pisseds
pisser
pisses
pissing
pissly
pissoff
pissoffed
pissoffer
pissoffes
pissoffing
pissoffly
pissoffs
pisss
polack
polacked
polacker
polackes
polacking
polackly
polacks
pollock
pollocked
pollocker
pollockes
pollocking
pollockly
pollocks
poon
pooned
pooner
poones
pooning
poonly
poons
poontang
poontanged
poontanger
poontanges
poontanging
poontangly
poontangs
porn
porned
porner
pornes
porning
pornly
porno
pornoed
pornoer
pornoes
pornography
pornographyed
pornographyer
pornographyes
pornographying
pornographyly
pornographys
pornoing
pornoly
pornos
porns
prick
pricked
pricker
prickes
pricking
prickly
pricks
prig
priged
priger
priges
priging
prigly
prigs
prostitute
prostituteed
prostituteer
prostitutees
prostituteing
prostitutely
prostitutes
prude
prudeed
prudeer
prudees
prudeing
prudely
prudes
punkass
punkassed
punkasser
punkasses
punkassing
punkassly
punkasss
punky
punkyed
punkyer
punkyes
punkying
punkyly
punkys
puss
pussed
pusser
pusses
pussies
pussiesed
pussieser
pussieses
pussiesing
pussiesly
pussiess
pussing
pussly
pusss
pussy
pussyed
pussyer
pussyes
pussying
pussyly
pussypounder
pussypoundered
pussypounderer
pussypounderes
pussypoundering
pussypounderly
pussypounders
pussys
puto
putoed
putoer
putoes
putoing
putoly
putos
queaf
queafed
queafer
queafes
queafing
queafly
queafs
queef
queefed
queefer
queefes
queefing
queefly
queefs
queer
queered
queerer
queeres
queering
queerly
queero
queeroed
queeroer
queeroes
queeroing
queeroly
queeros
queers
queersed
queerser
queerses
queersing
queersly
queerss
quicky
quickyed
quickyer
quickyes
quickying
quickyly
quickys
quim
quimed
quimer
quimes
quiming
quimly
quims
racy
racyed
racyer
racyes
racying
racyly
racys
rape
raped
rapeded
rapeder
rapedes
rapeding
rapedly
rapeds
rapeed
rapeer
rapees
rapeing
rapely
raper
rapered
raperer
raperes
rapering
raperly
rapers
rapes
rapist
rapisted
rapister
rapistes
rapisting
rapistly
rapists
raunch
raunched
rauncher
raunches
raunching
raunchly
raunchs
rectus
rectused
rectuser
rectuses
rectusing
rectusly
rectuss
reefer
reefered
reeferer
reeferes
reefering
reeferly
reefers
reetard
reetarded
reetarder
reetardes
reetarding
reetardly
reetards
reich
reiched
reicher
reiches
reiching
reichly
reichs
retard
retarded
retardeded
retardeder
retardedes
retardeding
retardedly
retardeds
retarder
retardes
retarding
retardly
retards
rimjob
rimjobed
rimjober
rimjobes
rimjobing
rimjobly
rimjobs
ritard
ritarded
ritarder
ritardes
ritarding
ritardly
ritards
rtard
rtarded
rtarder
rtardes
rtarding
rtardly
rtards
rum
rumed
rumer
rumes
ruming
rumly
rump
rumped
rumper
rumpes
rumping
rumply
rumprammer
rumprammered
rumprammerer
rumprammeres
rumprammering
rumprammerly
rumprammers
rumps
rums
ruski
ruskied
ruskier
ruskies
ruskiing
ruskily
ruskis
sadism
sadismed
sadismer
sadismes
sadisming
sadismly
sadisms
sadist
sadisted
sadister
sadistes
sadisting
sadistly
sadists
scag
scaged
scager
scages
scaging
scagly
scags
scantily
scantilyed
scantilyer
scantilyes
scantilying
scantilyly
scantilys
schlong
schlonged
schlonger
schlonges
schlonging
schlongly
schlongs
scrog
scroged
scroger
scroges
scroging
scrogly
scrogs
scrot
scrote
scroted
scroteed
scroteer
scrotees
scroteing
scrotely
scroter
scrotes
scroting
scrotly
scrots
scrotum
scrotumed
scrotumer
scrotumes
scrotuming
scrotumly
scrotums
scrud
scruded
scruder
scrudes
scruding
scrudly
scruds
scum
scumed
scumer
scumes
scuming
scumly
scums
seaman
seamaned
seamaner
seamanes
seamaning
seamanly
seamans
seamen
seamened
seamener
seamenes
seamening
seamenly
seamens
seduceed
seduceer
seducees
seduceing
seducely
seduces
semen
semened
semener
semenes
semening
semenly
semens
shamedame
shamedameed
shamedameer
shamedamees
shamedameing
shamedamely
shamedames
shit
shite
shiteater
shiteatered
shiteaterer
shiteateres
shiteatering
shiteaterly
shiteaters
shited
shiteed
shiteer
shitees
shiteing
shitely
shiter
shites
shitface
shitfaceed
shitfaceer
shitfacees
shitfaceing
shitfacely
shitfaces
shithead
shitheaded
shitheader
shitheades
shitheading
shitheadly
shitheads
shithole
shitholeed
shitholeer
shitholees
shitholeing
shitholely
shitholes
shithouse
shithouseed
shithouseer
shithousees
shithouseing
shithousely
shithouses
shiting
shitly
shits
shitsed
shitser
shitses
shitsing
shitsly
shitss
shitt
shitted
shitteded
shitteder
shittedes
shitteding
shittedly
shitteds
shitter
shittered
shitterer
shitteres
shittering
shitterly
shitters
shittes
shitting
shittly
shitts
shitty
shittyed
shittyer
shittyes
shittying
shittyly
shittys
shiz
shized
shizer
shizes
shizing
shizly
shizs
shooted
shooter
shootes
shooting
shootly
shoots
sissy
sissyed
sissyer
sissyes
sissying
sissyly
sissys
skag
skaged
skager
skages
skaging
skagly
skags
skank
skanked
skanker
skankes
skanking
skankly
skanks
slave
slaveed
slaveer
slavees
slaveing
slavely
slaves
sleaze
sleazeed
sleazeer
sleazees
sleazeing
sleazely
sleazes
sleazy
sleazyed
sleazyer
sleazyes
sleazying
sleazyly
sleazys
slut
slutdumper
slutdumpered
slutdumperer
slutdumperes
slutdumpering
slutdumperly
slutdumpers
sluted
sluter
slutes
sluting
slutkiss
slutkissed
slutkisser
slutkisses
slutkissing
slutkissly
slutkisss
slutly
sluts
slutsed
slutser
slutses
slutsing
slutsly
slutss
smegma
smegmaed
smegmaer
smegmaes
smegmaing
smegmaly
smegmas
smut
smuted
smuter
smutes
smuting
smutly
smuts
smutty
smuttyed
smuttyer
smuttyes
smuttying
smuttyly
smuttys
snatch
snatched
snatcher
snatches
snatching
snatchly
snatchs
sniper
snipered
sniperer
sniperes
snipering
sniperly
snipers
snort
snorted
snorter
snortes
snorting
snortly
snorts
snuff
snuffed
snuffer
snuffes
snuffing
snuffly
snuffs
sodom
sodomed
sodomer
sodomes
sodoming
sodomly
sodoms
spic
spiced
spicer
spices
spicing
spick
spicked
spicker
spickes
spicking
spickly
spicks
spicly
spics
spik
spoof
spoofed
spoofer
spoofes
spoofing
spoofly
spoofs
spooge
spoogeed
spoogeer
spoogees
spoogeing
spoogely
spooges
spunk
spunked
spunker
spunkes
spunking
spunkly
spunks
steamyed
steamyer
steamyes
steamying
steamyly
steamys
stfu
stfued
stfuer
stfues
stfuing
stfuly
stfus
stiffy
stiffyed
stiffyer
stiffyes
stiffying
stiffyly
stiffys
stoneded
stoneder
stonedes
stoneding
stonedly
stoneds
stupided
stupider
stupides
stupiding
stupidly
stupids
suckeded
suckeder
suckedes
suckeding
suckedly
suckeds
sucker
suckes
sucking
suckinged
suckinger
suckinges
suckinging
suckingly
suckings
suckly
sucks
sumofabiatch
sumofabiatched
sumofabiatcher
sumofabiatches
sumofabiatching
sumofabiatchly
sumofabiatchs
tard
tarded
tarder
tardes
tarding
tardly
tards
tawdry
tawdryed
tawdryer
tawdryes
tawdrying
tawdryly
tawdrys
teabagging
teabagginged
teabagginger
teabagginges
teabagginging
teabaggingly
teabaggings
terd
terded
terder
terdes
terding
terdly
terds
teste
testee
testeed
testeeed
testeeer
testeees
testeeing
testeely
testeer
testees
testeing
testely
testes
testesed
testeser
testeses
testesing
testesly
testess
testicle
testicleed
testicleer
testiclees
testicleing
testiclely
testicles
testis
testised
testiser
testises
testising
testisly
testiss
thrusted
thruster
thrustes
thrusting
thrustly
thrusts
thug
thuged
thuger
thuges
thuging
thugly
thugs
tinkle
tinkleed
tinkleer
tinklees
tinkleing
tinklely
tinkles
tit
tited
titer
tites
titfuck
titfucked
titfucker
titfuckes
titfucking
titfuckly
titfucks
titi
titied
titier
tities
titiing
titily
titing
titis
titly
tits
titsed
titser
titses
titsing
titsly
titss
tittiefucker
tittiefuckered
tittiefuckerer
tittiefuckeres
tittiefuckering
tittiefuckerly
tittiefuckers
titties
tittiesed
tittieser
tittieses
tittiesing
tittiesly
tittiess
titty
tittyed
tittyer
tittyes
tittyfuck
tittyfucked
tittyfucker
tittyfuckered
tittyfuckerer
tittyfuckeres
tittyfuckering
tittyfuckerly
tittyfuckers
tittyfuckes
tittyfucking
tittyfuckly
tittyfucks
tittying
tittyly
tittys
toke
tokeed
tokeer
tokees
tokeing
tokely
tokes
toots
tootsed
tootser
tootses
tootsing
tootsly
tootss
tramp
tramped
tramper
trampes
tramping
tramply
tramps
transsexualed
transsexualer
transsexuales
transsexualing
transsexually
transsexuals
trashy
trashyed
trashyer
trashyes
trashying
trashyly
trashys
tubgirl
tubgirled
tubgirler
tubgirles
tubgirling
tubgirlly
tubgirls
turd
turded
turder
turdes
turding
turdly
turds
tush
tushed
tusher
tushes
tushing
tushly
tushs
twat
twated
twater
twates
twating
twatly
twats
twatsed
twatser
twatses
twatsing
twatsly
twatss
undies
undiesed
undieser
undieses
undiesing
undiesly
undiess
unweded
unweder
unwedes
unweding
unwedly
unweds
uzi
uzied
uzier
uzies
uziing
uzily
uzis
vag
vaged
vager
vages
vaging
vagly
vags
valium
valiumed
valiumer
valiumes
valiuming
valiumly
valiums
venous
virgined
virginer
virgines
virgining
virginly
virgins
vixen
vixened
vixener
vixenes
vixening
vixenly
vixens
vodkaed
vodkaer
vodkaes
vodkaing
vodkaly
vodkas
voyeur
voyeured
voyeurer
voyeures
voyeuring
voyeurly
voyeurs
vulgar
vulgared
vulgarer
vulgares
vulgaring
vulgarly
vulgars
wang
wanged
wanger
wanges
wanging
wangly
wangs
wank
wanked
wanker
wankered
wankerer
wankeres
wankering
wankerly
wankers
wankes
wanking
wankly
wanks
wazoo
wazooed
wazooer
wazooes
wazooing
wazooly
wazoos
wedgie
wedgieed
wedgieer
wedgiees
wedgieing
wedgiely
wedgies
weeded
weeder
weedes
weeding
weedly
weeds
weenie
weenieed
weenieer
weeniees
weenieing
weeniely
weenies
weewee
weeweeed
weeweeer
weeweees
weeweeing
weeweely
weewees
weiner
weinered
weinerer
weineres
weinering
weinerly
weiners
weirdo
weirdoed
weirdoer
weirdoes
weirdoing
weirdoly
weirdos
wench
wenched
wencher
wenches
wenching
wenchly
wenchs
wetback
wetbacked
wetbacker
wetbackes
wetbacking
wetbackly
wetbacks
whitey
whiteyed
whiteyer
whiteyes
whiteying
whiteyly
whiteys
whiz
whized
whizer
whizes
whizing
whizly
whizs
whoralicious
whoralicioused
whoraliciouser
whoraliciouses
whoraliciousing
whoraliciously
whoraliciouss
whore
whorealicious
whorealicioused
whorealiciouser
whorealiciouses
whorealiciousing
whorealiciously
whorealiciouss
whored
whoreded
whoreder
whoredes
whoreding
whoredly
whoreds
whoreed
whoreer
whorees
whoreface
whorefaceed
whorefaceer
whorefacees
whorefaceing
whorefacely
whorefaces
whorehopper
whorehoppered
whorehopperer
whorehopperes
whorehoppering
whorehopperly
whorehoppers
whorehouse
whorehouseed
whorehouseer
whorehousees
whorehouseing
whorehousely
whorehouses
whoreing
whorely
whores
whoresed
whoreser
whoreses
whoresing
whoresly
whoress
whoring
whoringed
whoringer
whoringes
whoringing
whoringly
whorings
wigger
wiggered
wiggerer
wiggeres
wiggering
wiggerly
wiggers
woody
woodyed
woodyer
woodyes
woodying
woodyly
woodys
wop
woped
woper
wopes
woping
woply
wops
wtf
wtfed
wtfer
wtfes
wtfing
wtfly
wtfs
xxx
xxxed
xxxer
xxxes
xxxing
xxxly
xxxs
yeasty
yeastyed
yeastyer
yeastyes
yeastying
yeastyly
yeastys
yobbo
yobboed
yobboer
yobboes
yobboing
yobboly
yobbos
zoophile
zoophileed
zoophileer
zoophilees
zoophileing
zoophilely
zoophiles
anal
ass
ass lick
balls
ballsac
bisexual
bleach
causas
cheap
cost of miracles
cunt
display network stats
fart
fda and death
fda AND warn
fda AND warning
fda AND warns
feom
fuck
gfc
humira AND expensive
illegal
madvocate
masturbation
nuccitelli
overdose
porn
shit
snort
texarkana
direct\-acting antivirals
assistance
ombitasvir
support path
harvoni
abbvie
direct-acting antivirals
paritaprevir
advocacy
ledipasvir
vpak
ritonavir with dasabuvir
program
gilead
greedy
financial
needy
fake-ovir
viekira pak
v pak
sofosbuvir
support
oasis
discount
dasabuvir
protest
ritonavir
section[contains(@class, 'nav-hidden')]
footer[@id='footer']
div[contains(@class, 'pane-pub-article-cleveland-clinic')]
div[contains(@class, 'pane-pub-home-cleveland-clinic')]
div[contains(@class, 'pane-pub-topic-cleveland-clinic')]
div[contains(@class, 'panel-panel-inner')]
div[contains(@class, 'pane-node-field-article-topics')]
section[contains(@class, 'footer-nav-section-wrapper')]
Pinacidil induces vascular dilation and hyperemia in vivo and does not impact biophysical properties of neurons and astrocytes in vitro
Interactions between the brain and blood are essential to health. Metabolic supply of the brain is provided through the vasculature, and disruptions of this relationship, in extreme cases such as stroke, is a key characteristic of neurologic disease. Neuro-hemodynamic coupling is also demonstrated in healthy individuals on faster time scales in functional hyperemia, the local increase in blood flow and volume that accompanies neural activity.1,2
We have recently proposed a further level of interdependence between the two systems —ie, the hemoneural hypothesis—which predicts that hemodynamic events such as functional hyperemia will modulate neural activity.3 An impact of hemodynamics on neurons could occur through a number of mechanisms, including the activation of mechanoreceptors on astrocytes or neurons, a thermal impact of increased blood flow on ion channels and vesicle release, and the local increase and diffusion of blood-borne factors such as nitric oxide.4,5 Astrocytes are predicted to play a key role in hemo-neural modulation, as they are tightly coupled to the vascular system and participate in a number of neural functions.6,7 Through these mechanisms and others, hemodynamics could shift the “state” of the local neural circuit, thereby impacting information processing. This regulation of neural dynamics could also provide a homeostatic mechanism for promoting healthy brain function (eg, prevention of kindling).
To study the impact of hyperemia on neural and astrocytic activity in vivo, it is essential to independently control blood flow in the brain with means that do not directly impact neurons or astrocytes. Pinacidil is a sulfonylurea receptor agonist that opens the SUR2B potassium-sensitive ATP channel.8 In the telencephalon, SUR1 receptors are localized to neurons and glia.9,10 In contrast, SUR2 receptors are localized to vasculature, with SUR2A in cardiac and skeletal muscle, and SUR2B in vascular smooth muscle, with primary expression in smaller arteries, arterioles, and capillaries.11 By opening the SUR2B channel, pinacidil hyperpolarizes and relaxes smooth muscle, causing vasodilation. Pinacidil is a potent and selective SUR2B agonist, with a dissociation constant of 135 nM and a half maximal effective concentation (EC50) value of 680 nM.12 This agonist is approximately 5 times more specific for SUR2B than for SUR2A and shows approximately 5 orders of magnitude lower affinity for SUR1 (in the mM range).12–15 Previous studies have demonstrated the efficacy of this agent as a vasodilator.16–18
In the present study, we systematically examined the utility of pinacidil for the selective induction of hyperemia. First, we quantified the vasodilation induced by pinacidil in vivo, and examined local increases in blood volume in the parenchyma. These studies were conducted in anesthetized rats and awake mice. Second, we used in vitro slice recordings to examine whether direct application of relatively high concentrations of pinacidil would have any impact on the physiology of neurons and astrocytes. We found that (1) in vivo, pinacidil induces a level of vasodilation and increased local blood volume consistent with natural functional hyperemia across a variety of preparations, and (2) in vitro, pinacidil has no detectable impact on intrinsic biophysical measures in neurons and astrocytes.
METHODS
Animal preparation in vivo
To probe the impact of pinacidil on arterial diameter and parenchymal blood volume in vivo, we measured the effects of topical application to the primary somatosensory cortex (SI) of rats and mice. Sprague-Dawley rats (250–500 g) and C57BL/6 mice (~25 g) were anaesthetized with pentobarbital (50 mg/kg intraperitoneally initial dose, followed by 5-mg supplements as needed for maintenance). Animals were maintained at approximately 37°C by a heating blanket. Craniotomy (diameter of ~2 mm in rats, ~1 mm in mice) and durotomy were performed over SI, and the cortex was protected with Kwik-Cast silicone elastomer sealant (WPI, Sarasota, FL) while an imaging chamber was attached with dental cement. Kwik-Cast was removed, and the chamber filled with 0.9% saline and sealed with a round cover glass (avoiding bubbles) secured with cyanoacrylate.
Controlling visualization during drug delivery in vivo
To minimize brain motion and flow artifacts during visualization of hemodynamics in the rat preparation, we constructed a customized pressurized chamber with inflow and outflow for constant perfusion. The volume of the chamber was approximately 0.3 mL, and the flow through the system averaged about 2 mL/min. The chamber consisted of a plastic ring 1 cm in diameter and 3 mm high with a flat-top profile and a base shaped to the angle of the lateral skull edge over SI. In the wall of this chamber, three large holes were drilled and patched with pieces cut from rubber NMR septa (VWR International, West Chester, PA) to create resealable ports for drug application and bubble removal. Three additional permanent holes were drilled in the chamber walls, through which blunted 1-cm lengths of 18-gauge stainless steel needles were wedged and affixed with Super glue: one for artificial cerebrospinal fluid (ACSF) inflow, one for combined outflow, and the third for pressure regulation. The overall pressure of the chamber was regulated by a small vertical tube whose height (and thus fluid level) could be adjusted on a manipulator stand, and whose other end was open to the atmosphere. Inflow and outflow were controlled via regulators on a gravity feed system. In the mouse preparation, the need to control visualization was addressed by maintaining a constant rate of wicking in a smaller-profile open chamber, and a microfluidic switch with 12 μL of dead space was added to minimize propulsive impact and delay due to switching between solutions. Drug and ethanol solutions were delivered to rat and mouse chambers after being heated to physiological temperature (37°C).
Optical measurement of hemodynamics in vivo
We used a charge-coupled device camera (the Roper 512B, Princeton Instruments, Trenton, NJ) to image the cortical surface at a frame rate of approximately 4 Hz, with illumination from a voltage-regulated xenon arc lamp. A green band-pass filter (550 nm) was used to maximize imaging near the isosbestic point of hemoglobin, providing optimal vessel contrast and a surrogate measure for blood volume change in the parenchyma. Lenses (50 and 125 mm) were arranged in series to form a macroscope.
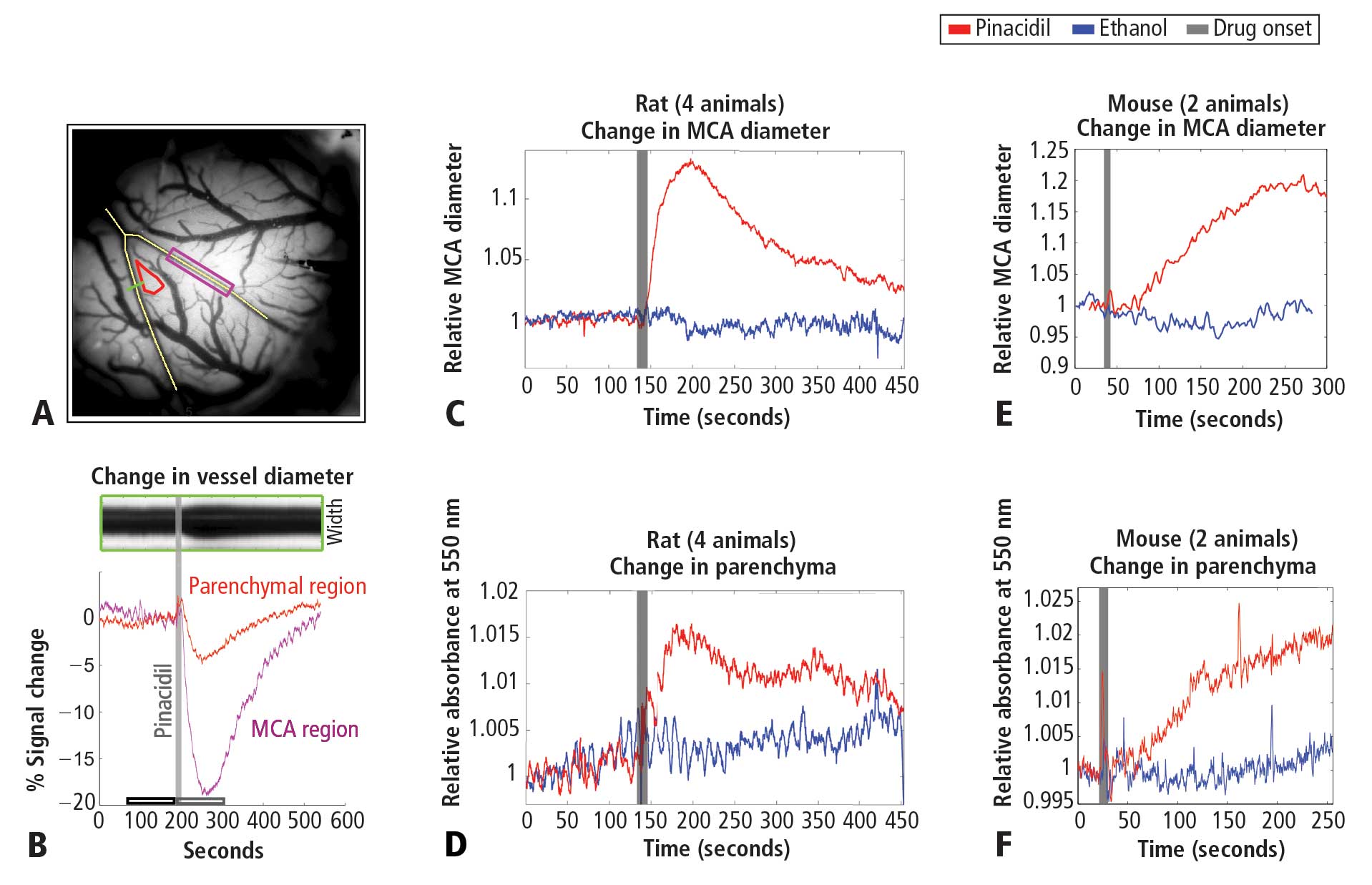
Pinacidil administration in vivo
Pinacidil is hydrophobic and was therefore dissolved in ethanol at approximately 12 mg/kg and then diluted 1:100 in ACSF to achieve a 400-μM solution in 1% ethanol. Stock solutions were stored at −20°C and diluted in fresh ACSF for each experiment. For each run, the cortex was imaged for 3 minutes to establish baseline. For the pressurized rat chamber, at the end of the baseline period, 0.1 to 0.3 mL of 400 μM pinacidil in 1% ethanol in ACSF or saline would be pumped into the 0.3-mL chamber (taking about 1 second). Simultaneously, an equivalent volume was drawn out to balance pressure by a push-pull pump with access through two of the resealable rubber ports.
Pinacidil administration in vitro
Coronal slices were prepared from Sprague-Dawley rats at postnatal day 14 to 40 and maintained in a submersion chamber at 27°C for recording. Solutions were prepared in ACSF: 125 mM NaCl, 2.5 mM KCl, 1 mM MgCl2, 1.25 mM NaH2PO4, 2 mM CaCl2, 25 mM NaHCO3, and 25 mM d-glucose). The applied solutions were 1% ethanol, ~400 μM pinacidil in 1% ethanol, and ACSF, all perfused with 5% CO2 in 95% O2 (carbogen).
For astrocyte recordings, slices were incubated immediately after cutting for 20 minutes in ACSF containing 50 μM sulforhodamine 101 (SR101), a water-soluble fluorescent dye specifically taken up by astrocytes.19 The slices were then allowed to rest for an hour before recording as usual. For fluorescence, the light source was a 100-W mercury arc lamp, with excitation and barrier filters and dichroic mirrors tailored to the spectral characteristics of SR101 (excitation ~586 nm, emission ~605 nm)
Slices were imaged under differential interference contrast optics with infrared illumination. Cells in layers 2/3 and in the same field and plane of view as a blood vessel greater than 20 μm in diameter were targeted, and vessel expansion was monitored during intracellular recording at approximately 4 Hz with a cooled charge-coupled device camera (Retiga EX, QImaging, Surrey, BC, Canada) connected to the microscope via a parfocal C-mount. This configuration also enabled imaging of neurons and astrocytes in the slice.
Drug/control solutions were switched every 90 to 180 seconds. Recording pipettes were filled with 120 mM KGlu, 10 mM NaCl, 20 mM KCl, 10 mM HEPES, 2mM Mg-ATP, 0.3 mM Na-GTP, 0.5 mM EGTA, and 0.3% to 1% biocytin (wt/vol) for subsequent visualization of the neurons.
RESULTS
Pinacidil induces vasodilation in anesthetized rats and awake mice
Figure 1A shows the cortical surface over SI. The green bisection line over the MCA shows the point of sampling for the darkness plot in the rectangular box in Figure 1B. The width of this dark band is the width of the MCA over time, showing expansion after pinacidil addition (gray bar). The point at which dilation is observed corresponds to a darkening in a parenchymal region (red triangle in Figure 1A) and over the MCA and surrounding cortex (purple rectangle in Figure 1A, reflecting expansion). Vasodilation and parenchymal signal increases were consistently observed on the first trial in all experiments (4 first runs from 4 anesthetized rats, Figure 1C and 1D). Dilation began less than 10 seconds after drug arrival, with maximal dilation and parenchymal darkening at an approximately 50- to 60-second latency. Ethanol in a 1:100 solution with ACSF under the same conditions evoked a nonsignificant reduction in vessel diameter and no change in parenchymal darkening. Following the first presentation, subsequent pinacidil effects were less reliable.
As shown in Figure 1E and 1F, the hemodynamic impact of pinacidil in anesthetized rats was replicated in awake, head-posted mice (2 mice, 2 runs each). Presentation of 220 or 440 μM pinacidil evoked comparable mean increases in arterial diameter (peak diameter increase of ~20%) and parenchymal darkening (peak increase of ~2%), effects that were repeatable within subjects in a single session (N = 2).
Pinacidil does not have direct effect on nonvascular tissue
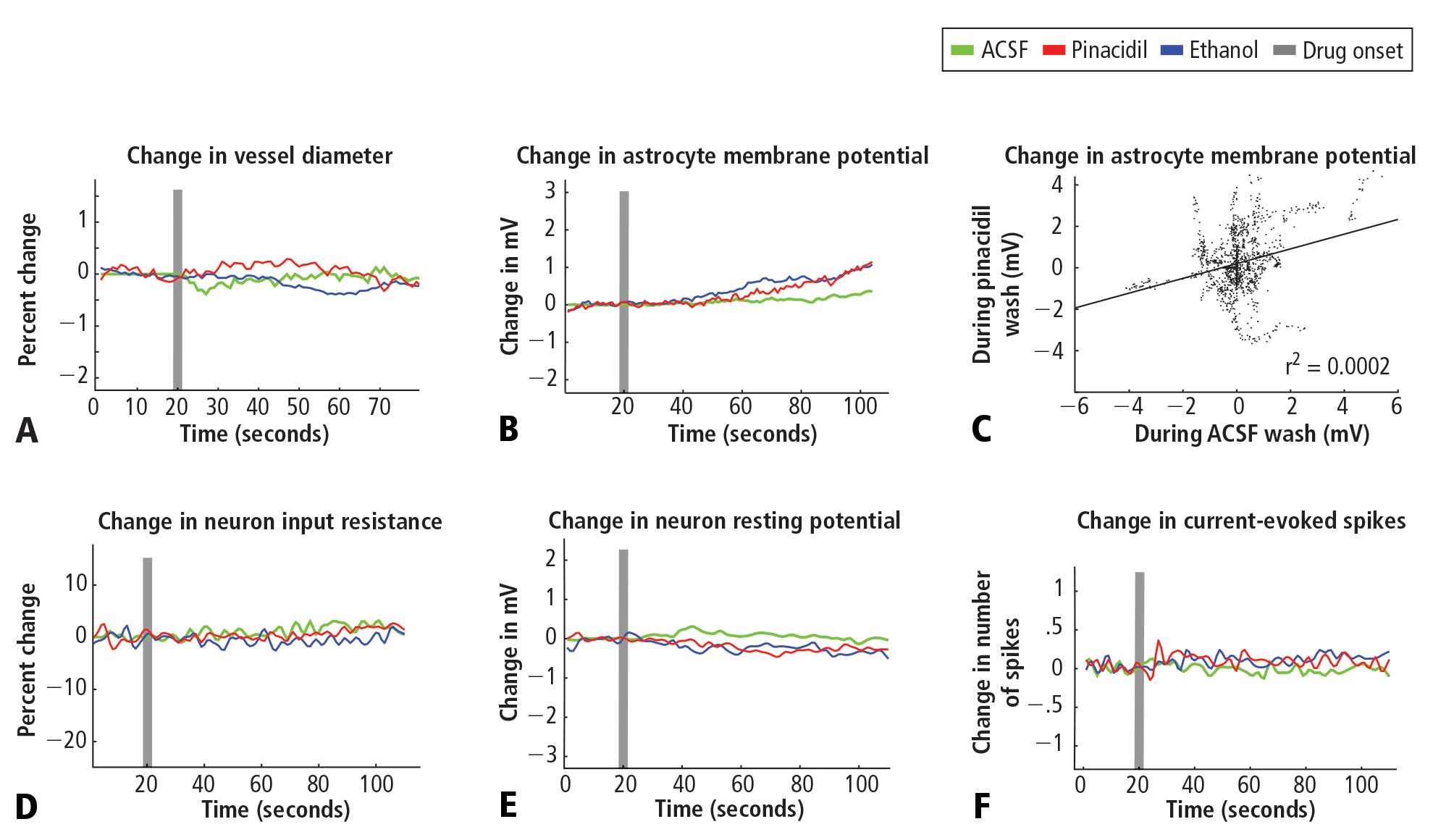
Pinacidil does not impact spiking probability or input resistance in neurons or membrane potential in neurons and astrocytes
In recordings from regular-spiking neurons of pyramidal shape (N = 30), we saw no change in any metric measured. At 50 seconds after application, approximately the time of peak vasodilatory effects in vivo, the resting membrane potential did not change (variation of −0.5 ± 0.9 mV standard deviation; Figure 2E), the spike rate induced by current injection did not change (variation of 0.1 ± 1.1 spikes/stimulation; Figure 2F), and input resistance did not change (variation of −0.7 ± 6.5 MΩ; Figure 2D). Similarly, in a limited subset of recordings from fast-spiking interneurons (N = 3), we did not observe any impact of pinacidil application. All significance tests were paired t tests (P > .10).
Astrocytes (N = 35) also showed no significant effects of pinacidil application, demonstrating only a slow depolarization during application of ethanol (1.3 ± 1.7 mV at 50 seconds after drug application) and pinacidil with ethanol (0.5 ± 2.1 mV) (Figure 2B). When we plotted the change in membrane potential at 10-second intervals since drug application (0 to 80 seconds post-drug), we found no trends in astrocytic response to either ACSF or pinacidil (Figure 2C).
During recordings at double our typical application dose (800 μM), we observed 2 pyramidal cells (out of 8) that showed depolarization (peak of 10 to 15 mV) and a loss of spike initiation capability. Following washout, these cells recovered membrane potential but spiking responses to current injection remained impaired. We did not evaluate the impact of this dose on vascular tone or rhythmic vasomotion.
In contrast to the absence of a detectable impact of pinacidil, we found that the membrane potentials of neurons and astrocytes were sensitive to flow rate. Decreasing flow rate caused a consistent depolarization of up to approximately 10 mV that showed an immediate onset, reaching a new baseline within 2 to 5 seconds; increasing flow rate had the opposite effect. In preliminary experiments, we observed two astrocytes that depolarized on switching to the pinacidil solution. These two recordings were obtained prior to placement of an inline pressure meter in the flow pathway that allowed us to monitor and exclude trials that showed flow changes. In the 35 subsequent recordings that did not have flow changes, we never observed a detectable impact of either ethanol or pinacidil on astroctyes or neurons. We also noted that neurons and astrocytes were more likely to die and/or to lose recording quality during a cycle of ethanol or pinacidil presentation, as opposed to ACSF presentation.
DISCUSSION
Pinacidil provides an effective means of inducing vasodilation in vivo. At concentrations less than 400 μM, pinacidil is also selective for cortical vascular smooth muscle, exhibiting no direct effect on intrinsic properties of neurons or astrocytes. As an independent means to induce increased vasodilation and blood volume in a manner analogous to that seen in functional hyperemia, pinacidil provides a viable method for testing the impact of hyperemic events on neural or astrocytic activity. Pinacidil may also be a selective means of emulating other normal hemodynamic phenomena and could have therapeutic applications, such as targeted administration of pinacidil in response to acute vessel obstruction to maintain sufficient perfusion.
The hemodynamic effects induced by pinacidil are similar to natural functional hyperemia. In SI during sensory stimulation in rodents, increases in total oxygenated hemoglobin during sensory stimulation— analogous to our measurement of cortical darkening at 550 nm—peak in a range of 2% to 5%,20–22 and arteries/arterioles dilate 10% to 20%.23 The time course of pinacidil’s effects also parallels the sustained response to continued sensory drive. Arterial diameter in rodent SI and the blood oxygen level–dependent response on functional magnetic resonance imaging in humans and rodents remain high when tactile input is sustained for periods lasting tens of seconds,23,24 as they do under pinacidil application.
Although pinacidil represents an important step forward in our ability to control blood flow while probing the impact of hemodynamics in cortex, it has limitations. The drug is only capable of producing vasodilation; drugs in the same family that block the SUR2B channels to create vasoconstriction (such as diazoxide or glibenclamide) or thromboxane receptor agonists18,25,26 are unfortunately known to be nonspecific, affecting neurons as well as blood vessels. Pinacidil is also not water-soluble, requiring its dissolution in ethanol or DMSO, agents that can have confounding impacts on the system. Applied in vivo, pinacidil also does not appear to wash out fully, or its impact on smooth muscles persists, so that the first trial in each animal is the most consistent and effective one. These limitations stated, this pharmacological approach nevertheless represents a unique means of selective hyperemia induction in vivo.
- Roy CS, Sherrington CS. On the regulation of the blood-supply of the brain. J Physiol 1890; 11:85–108,158-7–158-17.
- Raichle ME. Behind the scenes of functional brain imaging: a historical and physiological perspective. Proc Natl Acad Sci USA 1998; 95:765–772.
- Moore CI, Cao R. The hemo-neural hypothesis: on the role of blood flow in information processing. J Neurophysiol 2008; 99:2035–2047.
- Garthwaite G, Bartus K, Malcolm D, et al. Signaling from blood vessels to CNS axons through nitric oxide. J Neurosci 2006; 26:7730–7740.
- Kozlov AS, Angulo MC, Audinat E, Charpak S. Target cellspecific modulation of neuronal activity by astrocytes. Proc Natl Acad Sci USA 2006; 103:10058–10063.
- Nedergaard M, Ransom B, Goldman SA. New roles for astrocytes: redefining the functional architecture of the brain. Trends Neurosci 2003; 26:523–530.
- Haydon PG, Carmignoto G. Astrocyte control of synaptic transmission and neurovascular coupling. Physiol Rev 2006; 86:1009–1031.
- Ashcroft FM, Gribble FM. New windows on the mechanism of action of K(ATP) channel openers. Trends Pharmacol Sci 2000; 21:439–445.
- Levin BE, Dunn-Meynell AA, Routh VH. Brain glucosensing and the K(ATP) channel. Nat Neurosci 2001; 4:459–460.
- Zawar C, Plant TD, Schirra C, Konnerth A, Neumcke B. Celltype specific expression of ATP-sensitive potassium channels in the rat hippocampus. J Physiol 1999; 514:327–341.
- Li L, Wu J, Jiang C. Differential expression of Kir6.1 and SUR2B mRNAs in the vasculature of various tissues in rats. J Membr Biol 2003; 196:61–69.
- Schwanstecher M, Sieverding C, Dorschner H, et al. Potassium channel openers require ATP to bind to and act through sulfonylurea receptors. EMBO J 1998; 17:5529–5535.
- Shindo T, Yamada M, Isomoto S, Horio Y, Kurachi Y. SUR2 subtype (A and B)-dependent differential activation of the cloned ATP-sensitive K+ channels by pinacidil and nicorandil. Br J Pharmacol 1998; 124:985–991.
- Russ U, Lange U, Loffler-Walz C, Hambrock A, Quast U. Binding and effect of KATP channel openers in the absence of Mg2+. Br J Pharmacol 2003; 139:368–380.
- Higdon NR, Khan SA, Buchanan LV, Meisheri KD. Tissue and species variation in the vascular receptor binding of 3H-P1075, a potent KATP opener vasodilator. J Pharmacol Exp Ther 1997; 280:255–260.
- Wahl M. The effects of pinacidil and tolbutamide in feline pial arteries in situ. Pflugers Arch 1989; 415:250–252.
- Hempelmann RG, Barth HL, Mehdorn HM, Pradel RH, Ziegler A. Effects of potassium channel openers in isolated human cerebral arteries. Neurosurgery 1995; 37:1146–1153.
- Quayle JM, Nelson MT, Standen NB. ATP-sensitive and inwardly rectifying potassium channels in smooth muscle. Physiol Rev 1997; 77:1165–1232.
- Nimmerjahn A, Kirchhoff F, Kerr JN, Helmchen F. Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat Methods 2004; 1:31–37.
- Kong Y, Zheng Y, Johnston D, et al. A model of the dynamic relationship between blood flow and volume changes during brain activation. J Cereb Blood Flow Metab 2004; 24:1382–1392.
- Martin C, Martindale J, Berwick J, Mayhew J. Investigating neural-hemodynamic coupling and the hemodynamic response function in the awake rat. Neuroimage 2006; 32:33–48.
- Jones M, Devonshire IM, Berwick J, Martin C, Redgrave P, Mayhew J. Alterered neurovascular coupling during information-processing states. Eur J Neurosci 2008; 27:2758–2772.
- Woolsey TA, Rovainen CM, Cox SB, et al. Neuronal units linked to microvascular modules in cerebral cortex: response elements for imaging the brain. Cereb Cortex 1996; 6:647–660.
- Moore CI, Stern CE, Corkin S, et al. Segregation of somatosensory activation in the human rolandic cortex using fMRI. J Neurophysiol 2000; 84:558–569.
- Crépel V, Krnjević K, Ben-Ari Y. Sulphonylureas reduce the slowly inactivating D-type outward current in rat hippocampal neurons. J Physiol 1993; 466:39–54.
- Lovick TA, Brown LA, Key BJ. Neuronal activity-related coupling in cortical arterioles: involvement of astrocyte-derived factors. Exp Physiol 2005; 90:131–140.
Interactions between the brain and blood are essential to health. Metabolic supply of the brain is provided through the vasculature, and disruptions of this relationship, in extreme cases such as stroke, is a key characteristic of neurologic disease. Neuro-hemodynamic coupling is also demonstrated in healthy individuals on faster time scales in functional hyperemia, the local increase in blood flow and volume that accompanies neural activity.1,2
We have recently proposed a further level of interdependence between the two systems —ie, the hemoneural hypothesis—which predicts that hemodynamic events such as functional hyperemia will modulate neural activity.3 An impact of hemodynamics on neurons could occur through a number of mechanisms, including the activation of mechanoreceptors on astrocytes or neurons, a thermal impact of increased blood flow on ion channels and vesicle release, and the local increase and diffusion of blood-borne factors such as nitric oxide.4,5 Astrocytes are predicted to play a key role in hemo-neural modulation, as they are tightly coupled to the vascular system and participate in a number of neural functions.6,7 Through these mechanisms and others, hemodynamics could shift the “state” of the local neural circuit, thereby impacting information processing. This regulation of neural dynamics could also provide a homeostatic mechanism for promoting healthy brain function (eg, prevention of kindling).
To study the impact of hyperemia on neural and astrocytic activity in vivo, it is essential to independently control blood flow in the brain with means that do not directly impact neurons or astrocytes. Pinacidil is a sulfonylurea receptor agonist that opens the SUR2B potassium-sensitive ATP channel.8 In the telencephalon, SUR1 receptors are localized to neurons and glia.9,10 In contrast, SUR2 receptors are localized to vasculature, with SUR2A in cardiac and skeletal muscle, and SUR2B in vascular smooth muscle, with primary expression in smaller arteries, arterioles, and capillaries.11 By opening the SUR2B channel, pinacidil hyperpolarizes and relaxes smooth muscle, causing vasodilation. Pinacidil is a potent and selective SUR2B agonist, with a dissociation constant of 135 nM and a half maximal effective concentation (EC50) value of 680 nM.12 This agonist is approximately 5 times more specific for SUR2B than for SUR2A and shows approximately 5 orders of magnitude lower affinity for SUR1 (in the mM range).12–15 Previous studies have demonstrated the efficacy of this agent as a vasodilator.16–18
In the present study, we systematically examined the utility of pinacidil for the selective induction of hyperemia. First, we quantified the vasodilation induced by pinacidil in vivo, and examined local increases in blood volume in the parenchyma. These studies were conducted in anesthetized rats and awake mice. Second, we used in vitro slice recordings to examine whether direct application of relatively high concentrations of pinacidil would have any impact on the physiology of neurons and astrocytes. We found that (1) in vivo, pinacidil induces a level of vasodilation and increased local blood volume consistent with natural functional hyperemia across a variety of preparations, and (2) in vitro, pinacidil has no detectable impact on intrinsic biophysical measures in neurons and astrocytes.
METHODS
Animal preparation in vivo
To probe the impact of pinacidil on arterial diameter and parenchymal blood volume in vivo, we measured the effects of topical application to the primary somatosensory cortex (SI) of rats and mice. Sprague-Dawley rats (250–500 g) and C57BL/6 mice (~25 g) were anaesthetized with pentobarbital (50 mg/kg intraperitoneally initial dose, followed by 5-mg supplements as needed for maintenance). Animals were maintained at approximately 37°C by a heating blanket. Craniotomy (diameter of ~2 mm in rats, ~1 mm in mice) and durotomy were performed over SI, and the cortex was protected with Kwik-Cast silicone elastomer sealant (WPI, Sarasota, FL) while an imaging chamber was attached with dental cement. Kwik-Cast was removed, and the chamber filled with 0.9% saline and sealed with a round cover glass (avoiding bubbles) secured with cyanoacrylate.
Controlling visualization during drug delivery in vivo
To minimize brain motion and flow artifacts during visualization of hemodynamics in the rat preparation, we constructed a customized pressurized chamber with inflow and outflow for constant perfusion. The volume of the chamber was approximately 0.3 mL, and the flow through the system averaged about 2 mL/min. The chamber consisted of a plastic ring 1 cm in diameter and 3 mm high with a flat-top profile and a base shaped to the angle of the lateral skull edge over SI. In the wall of this chamber, three large holes were drilled and patched with pieces cut from rubber NMR septa (VWR International, West Chester, PA) to create resealable ports for drug application and bubble removal. Three additional permanent holes were drilled in the chamber walls, through which blunted 1-cm lengths of 18-gauge stainless steel needles were wedged and affixed with Super glue: one for artificial cerebrospinal fluid (ACSF) inflow, one for combined outflow, and the third for pressure regulation. The overall pressure of the chamber was regulated by a small vertical tube whose height (and thus fluid level) could be adjusted on a manipulator stand, and whose other end was open to the atmosphere. Inflow and outflow were controlled via regulators on a gravity feed system. In the mouse preparation, the need to control visualization was addressed by maintaining a constant rate of wicking in a smaller-profile open chamber, and a microfluidic switch with 12 μL of dead space was added to minimize propulsive impact and delay due to switching between solutions. Drug and ethanol solutions were delivered to rat and mouse chambers after being heated to physiological temperature (37°C).
Optical measurement of hemodynamics in vivo
We used a charge-coupled device camera (the Roper 512B, Princeton Instruments, Trenton, NJ) to image the cortical surface at a frame rate of approximately 4 Hz, with illumination from a voltage-regulated xenon arc lamp. A green band-pass filter (550 nm) was used to maximize imaging near the isosbestic point of hemoglobin, providing optimal vessel contrast and a surrogate measure for blood volume change in the parenchyma. Lenses (50 and 125 mm) were arranged in series to form a macroscope.
Pinacidil administration in vivo
Pinacidil is hydrophobic and was therefore dissolved in ethanol at approximately 12 mg/kg and then diluted 1:100 in ACSF to achieve a 400-μM solution in 1% ethanol. Stock solutions were stored at −20°C and diluted in fresh ACSF for each experiment. For each run, the cortex was imaged for 3 minutes to establish baseline. For the pressurized rat chamber, at the end of the baseline period, 0.1 to 0.3 mL of 400 μM pinacidil in 1% ethanol in ACSF or saline would be pumped into the 0.3-mL chamber (taking about 1 second). Simultaneously, an equivalent volume was drawn out to balance pressure by a push-pull pump with access through two of the resealable rubber ports.
Pinacidil administration in vitro
Coronal slices were prepared from Sprague-Dawley rats at postnatal day 14 to 40 and maintained in a submersion chamber at 27°C for recording. Solutions were prepared in ACSF: 125 mM NaCl, 2.5 mM KCl, 1 mM MgCl2, 1.25 mM NaH2PO4, 2 mM CaCl2, 25 mM NaHCO3, and 25 mM d-glucose). The applied solutions were 1% ethanol, ~400 μM pinacidil in 1% ethanol, and ACSF, all perfused with 5% CO2 in 95% O2 (carbogen).
For astrocyte recordings, slices were incubated immediately after cutting for 20 minutes in ACSF containing 50 μM sulforhodamine 101 (SR101), a water-soluble fluorescent dye specifically taken up by astrocytes.19 The slices were then allowed to rest for an hour before recording as usual. For fluorescence, the light source was a 100-W mercury arc lamp, with excitation and barrier filters and dichroic mirrors tailored to the spectral characteristics of SR101 (excitation ~586 nm, emission ~605 nm)
Slices were imaged under differential interference contrast optics with infrared illumination. Cells in layers 2/3 and in the same field and plane of view as a blood vessel greater than 20 μm in diameter were targeted, and vessel expansion was monitored during intracellular recording at approximately 4 Hz with a cooled charge-coupled device camera (Retiga EX, QImaging, Surrey, BC, Canada) connected to the microscope via a parfocal C-mount. This configuration also enabled imaging of neurons and astrocytes in the slice.
Drug/control solutions were switched every 90 to 180 seconds. Recording pipettes were filled with 120 mM KGlu, 10 mM NaCl, 20 mM KCl, 10 mM HEPES, 2mM Mg-ATP, 0.3 mM Na-GTP, 0.5 mM EGTA, and 0.3% to 1% biocytin (wt/vol) for subsequent visualization of the neurons.
RESULTS
Pinacidil induces vasodilation in anesthetized rats and awake mice
Figure 1A shows the cortical surface over SI. The green bisection line over the MCA shows the point of sampling for the darkness plot in the rectangular box in Figure 1B. The width of this dark band is the width of the MCA over time, showing expansion after pinacidil addition (gray bar). The point at which dilation is observed corresponds to a darkening in a parenchymal region (red triangle in Figure 1A) and over the MCA and surrounding cortex (purple rectangle in Figure 1A, reflecting expansion). Vasodilation and parenchymal signal increases were consistently observed on the first trial in all experiments (4 first runs from 4 anesthetized rats, Figure 1C and 1D). Dilation began less than 10 seconds after drug arrival, with maximal dilation and parenchymal darkening at an approximately 50- to 60-second latency. Ethanol in a 1:100 solution with ACSF under the same conditions evoked a nonsignificant reduction in vessel diameter and no change in parenchymal darkening. Following the first presentation, subsequent pinacidil effects were less reliable.
As shown in Figure 1E and 1F, the hemodynamic impact of pinacidil in anesthetized rats was replicated in awake, head-posted mice (2 mice, 2 runs each). Presentation of 220 or 440 μM pinacidil evoked comparable mean increases in arterial diameter (peak diameter increase of ~20%) and parenchymal darkening (peak increase of ~2%), effects that were repeatable within subjects in a single session (N = 2).
Pinacidil does not have direct effect on nonvascular tissue
Pinacidil does not impact spiking probability or input resistance in neurons or membrane potential in neurons and astrocytes
In recordings from regular-spiking neurons of pyramidal shape (N = 30), we saw no change in any metric measured. At 50 seconds after application, approximately the time of peak vasodilatory effects in vivo, the resting membrane potential did not change (variation of −0.5 ± 0.9 mV standard deviation; Figure 2E), the spike rate induced by current injection did not change (variation of 0.1 ± 1.1 spikes/stimulation; Figure 2F), and input resistance did not change (variation of −0.7 ± 6.5 MΩ; Figure 2D). Similarly, in a limited subset of recordings from fast-spiking interneurons (N = 3), we did not observe any impact of pinacidil application. All significance tests were paired t tests (P > .10).
Astrocytes (N = 35) also showed no significant effects of pinacidil application, demonstrating only a slow depolarization during application of ethanol (1.3 ± 1.7 mV at 50 seconds after drug application) and pinacidil with ethanol (0.5 ± 2.1 mV) (Figure 2B). When we plotted the change in membrane potential at 10-second intervals since drug application (0 to 80 seconds post-drug), we found no trends in astrocytic response to either ACSF or pinacidil (Figure 2C).
During recordings at double our typical application dose (800 μM), we observed 2 pyramidal cells (out of 8) that showed depolarization (peak of 10 to 15 mV) and a loss of spike initiation capability. Following washout, these cells recovered membrane potential but spiking responses to current injection remained impaired. We did not evaluate the impact of this dose on vascular tone or rhythmic vasomotion.
In contrast to the absence of a detectable impact of pinacidil, we found that the membrane potentials of neurons and astrocytes were sensitive to flow rate. Decreasing flow rate caused a consistent depolarization of up to approximately 10 mV that showed an immediate onset, reaching a new baseline within 2 to 5 seconds; increasing flow rate had the opposite effect. In preliminary experiments, we observed two astrocytes that depolarized on switching to the pinacidil solution. These two recordings were obtained prior to placement of an inline pressure meter in the flow pathway that allowed us to monitor and exclude trials that showed flow changes. In the 35 subsequent recordings that did not have flow changes, we never observed a detectable impact of either ethanol or pinacidil on astroctyes or neurons. We also noted that neurons and astrocytes were more likely to die and/or to lose recording quality during a cycle of ethanol or pinacidil presentation, as opposed to ACSF presentation.
DISCUSSION
Pinacidil provides an effective means of inducing vasodilation in vivo. At concentrations less than 400 μM, pinacidil is also selective for cortical vascular smooth muscle, exhibiting no direct effect on intrinsic properties of neurons or astrocytes. As an independent means to induce increased vasodilation and blood volume in a manner analogous to that seen in functional hyperemia, pinacidil provides a viable method for testing the impact of hyperemic events on neural or astrocytic activity. Pinacidil may also be a selective means of emulating other normal hemodynamic phenomena and could have therapeutic applications, such as targeted administration of pinacidil in response to acute vessel obstruction to maintain sufficient perfusion.
The hemodynamic effects induced by pinacidil are similar to natural functional hyperemia. In SI during sensory stimulation in rodents, increases in total oxygenated hemoglobin during sensory stimulation— analogous to our measurement of cortical darkening at 550 nm—peak in a range of 2% to 5%,20–22 and arteries/arterioles dilate 10% to 20%.23 The time course of pinacidil’s effects also parallels the sustained response to continued sensory drive. Arterial diameter in rodent SI and the blood oxygen level–dependent response on functional magnetic resonance imaging in humans and rodents remain high when tactile input is sustained for periods lasting tens of seconds,23,24 as they do under pinacidil application.
Although pinacidil represents an important step forward in our ability to control blood flow while probing the impact of hemodynamics in cortex, it has limitations. The drug is only capable of producing vasodilation; drugs in the same family that block the SUR2B channels to create vasoconstriction (such as diazoxide or glibenclamide) or thromboxane receptor agonists18,25,26 are unfortunately known to be nonspecific, affecting neurons as well as blood vessels. Pinacidil is also not water-soluble, requiring its dissolution in ethanol or DMSO, agents that can have confounding impacts on the system. Applied in vivo, pinacidil also does not appear to wash out fully, or its impact on smooth muscles persists, so that the first trial in each animal is the most consistent and effective one. These limitations stated, this pharmacological approach nevertheless represents a unique means of selective hyperemia induction in vivo.
Interactions between the brain and blood are essential to health. Metabolic supply of the brain is provided through the vasculature, and disruptions of this relationship, in extreme cases such as stroke, is a key characteristic of neurologic disease. Neuro-hemodynamic coupling is also demonstrated in healthy individuals on faster time scales in functional hyperemia, the local increase in blood flow and volume that accompanies neural activity.1,2
We have recently proposed a further level of interdependence between the two systems —ie, the hemoneural hypothesis—which predicts that hemodynamic events such as functional hyperemia will modulate neural activity.3 An impact of hemodynamics on neurons could occur through a number of mechanisms, including the activation of mechanoreceptors on astrocytes or neurons, a thermal impact of increased blood flow on ion channels and vesicle release, and the local increase and diffusion of blood-borne factors such as nitric oxide.4,5 Astrocytes are predicted to play a key role in hemo-neural modulation, as they are tightly coupled to the vascular system and participate in a number of neural functions.6,7 Through these mechanisms and others, hemodynamics could shift the “state” of the local neural circuit, thereby impacting information processing. This regulation of neural dynamics could also provide a homeostatic mechanism for promoting healthy brain function (eg, prevention of kindling).
To study the impact of hyperemia on neural and astrocytic activity in vivo, it is essential to independently control blood flow in the brain with means that do not directly impact neurons or astrocytes. Pinacidil is a sulfonylurea receptor agonist that opens the SUR2B potassium-sensitive ATP channel.8 In the telencephalon, SUR1 receptors are localized to neurons and glia.9,10 In contrast, SUR2 receptors are localized to vasculature, with SUR2A in cardiac and skeletal muscle, and SUR2B in vascular smooth muscle, with primary expression in smaller arteries, arterioles, and capillaries.11 By opening the SUR2B channel, pinacidil hyperpolarizes and relaxes smooth muscle, causing vasodilation. Pinacidil is a potent and selective SUR2B agonist, with a dissociation constant of 135 nM and a half maximal effective concentation (EC50) value of 680 nM.12 This agonist is approximately 5 times more specific for SUR2B than for SUR2A and shows approximately 5 orders of magnitude lower affinity for SUR1 (in the mM range).12–15 Previous studies have demonstrated the efficacy of this agent as a vasodilator.16–18
In the present study, we systematically examined the utility of pinacidil for the selective induction of hyperemia. First, we quantified the vasodilation induced by pinacidil in vivo, and examined local increases in blood volume in the parenchyma. These studies were conducted in anesthetized rats and awake mice. Second, we used in vitro slice recordings to examine whether direct application of relatively high concentrations of pinacidil would have any impact on the physiology of neurons and astrocytes. We found that (1) in vivo, pinacidil induces a level of vasodilation and increased local blood volume consistent with natural functional hyperemia across a variety of preparations, and (2) in vitro, pinacidil has no detectable impact on intrinsic biophysical measures in neurons and astrocytes.
METHODS
Animal preparation in vivo
To probe the impact of pinacidil on arterial diameter and parenchymal blood volume in vivo, we measured the effects of topical application to the primary somatosensory cortex (SI) of rats and mice. Sprague-Dawley rats (250–500 g) and C57BL/6 mice (~25 g) were anaesthetized with pentobarbital (50 mg/kg intraperitoneally initial dose, followed by 5-mg supplements as needed for maintenance). Animals were maintained at approximately 37°C by a heating blanket. Craniotomy (diameter of ~2 mm in rats, ~1 mm in mice) and durotomy were performed over SI, and the cortex was protected with Kwik-Cast silicone elastomer sealant (WPI, Sarasota, FL) while an imaging chamber was attached with dental cement. Kwik-Cast was removed, and the chamber filled with 0.9% saline and sealed with a round cover glass (avoiding bubbles) secured with cyanoacrylate.
Controlling visualization during drug delivery in vivo
To minimize brain motion and flow artifacts during visualization of hemodynamics in the rat preparation, we constructed a customized pressurized chamber with inflow and outflow for constant perfusion. The volume of the chamber was approximately 0.3 mL, and the flow through the system averaged about 2 mL/min. The chamber consisted of a plastic ring 1 cm in diameter and 3 mm high with a flat-top profile and a base shaped to the angle of the lateral skull edge over SI. In the wall of this chamber, three large holes were drilled and patched with pieces cut from rubber NMR septa (VWR International, West Chester, PA) to create resealable ports for drug application and bubble removal. Three additional permanent holes were drilled in the chamber walls, through which blunted 1-cm lengths of 18-gauge stainless steel needles were wedged and affixed with Super glue: one for artificial cerebrospinal fluid (ACSF) inflow, one for combined outflow, and the third for pressure regulation. The overall pressure of the chamber was regulated by a small vertical tube whose height (and thus fluid level) could be adjusted on a manipulator stand, and whose other end was open to the atmosphere. Inflow and outflow were controlled via regulators on a gravity feed system. In the mouse preparation, the need to control visualization was addressed by maintaining a constant rate of wicking in a smaller-profile open chamber, and a microfluidic switch with 12 μL of dead space was added to minimize propulsive impact and delay due to switching between solutions. Drug and ethanol solutions were delivered to rat and mouse chambers after being heated to physiological temperature (37°C).
Optical measurement of hemodynamics in vivo
We used a charge-coupled device camera (the Roper 512B, Princeton Instruments, Trenton, NJ) to image the cortical surface at a frame rate of approximately 4 Hz, with illumination from a voltage-regulated xenon arc lamp. A green band-pass filter (550 nm) was used to maximize imaging near the isosbestic point of hemoglobin, providing optimal vessel contrast and a surrogate measure for blood volume change in the parenchyma. Lenses (50 and 125 mm) were arranged in series to form a macroscope.
Pinacidil administration in vivo
Pinacidil is hydrophobic and was therefore dissolved in ethanol at approximately 12 mg/kg and then diluted 1:100 in ACSF to achieve a 400-μM solution in 1% ethanol. Stock solutions were stored at −20°C and diluted in fresh ACSF for each experiment. For each run, the cortex was imaged for 3 minutes to establish baseline. For the pressurized rat chamber, at the end of the baseline period, 0.1 to 0.3 mL of 400 μM pinacidil in 1% ethanol in ACSF or saline would be pumped into the 0.3-mL chamber (taking about 1 second). Simultaneously, an equivalent volume was drawn out to balance pressure by a push-pull pump with access through two of the resealable rubber ports.
Pinacidil administration in vitro
Coronal slices were prepared from Sprague-Dawley rats at postnatal day 14 to 40 and maintained in a submersion chamber at 27°C for recording. Solutions were prepared in ACSF: 125 mM NaCl, 2.5 mM KCl, 1 mM MgCl2, 1.25 mM NaH2PO4, 2 mM CaCl2, 25 mM NaHCO3, and 25 mM d-glucose). The applied solutions were 1% ethanol, ~400 μM pinacidil in 1% ethanol, and ACSF, all perfused with 5% CO2 in 95% O2 (carbogen).
For astrocyte recordings, slices were incubated immediately after cutting for 20 minutes in ACSF containing 50 μM sulforhodamine 101 (SR101), a water-soluble fluorescent dye specifically taken up by astrocytes.19 The slices were then allowed to rest for an hour before recording as usual. For fluorescence, the light source was a 100-W mercury arc lamp, with excitation and barrier filters and dichroic mirrors tailored to the spectral characteristics of SR101 (excitation ~586 nm, emission ~605 nm)
Slices were imaged under differential interference contrast optics with infrared illumination. Cells in layers 2/3 and in the same field and plane of view as a blood vessel greater than 20 μm in diameter were targeted, and vessel expansion was monitored during intracellular recording at approximately 4 Hz with a cooled charge-coupled device camera (Retiga EX, QImaging, Surrey, BC, Canada) connected to the microscope via a parfocal C-mount. This configuration also enabled imaging of neurons and astrocytes in the slice.
Drug/control solutions were switched every 90 to 180 seconds. Recording pipettes were filled with 120 mM KGlu, 10 mM NaCl, 20 mM KCl, 10 mM HEPES, 2mM Mg-ATP, 0.3 mM Na-GTP, 0.5 mM EGTA, and 0.3% to 1% biocytin (wt/vol) for subsequent visualization of the neurons.
RESULTS
Pinacidil induces vasodilation in anesthetized rats and awake mice
Figure 1A shows the cortical surface over SI. The green bisection line over the MCA shows the point of sampling for the darkness plot in the rectangular box in Figure 1B. The width of this dark band is the width of the MCA over time, showing expansion after pinacidil addition (gray bar). The point at which dilation is observed corresponds to a darkening in a parenchymal region (red triangle in Figure 1A) and over the MCA and surrounding cortex (purple rectangle in Figure 1A, reflecting expansion). Vasodilation and parenchymal signal increases were consistently observed on the first trial in all experiments (4 first runs from 4 anesthetized rats, Figure 1C and 1D). Dilation began less than 10 seconds after drug arrival, with maximal dilation and parenchymal darkening at an approximately 50- to 60-second latency. Ethanol in a 1:100 solution with ACSF under the same conditions evoked a nonsignificant reduction in vessel diameter and no change in parenchymal darkening. Following the first presentation, subsequent pinacidil effects were less reliable.
As shown in Figure 1E and 1F, the hemodynamic impact of pinacidil in anesthetized rats was replicated in awake, head-posted mice (2 mice, 2 runs each). Presentation of 220 or 440 μM pinacidil evoked comparable mean increases in arterial diameter (peak diameter increase of ~20%) and parenchymal darkening (peak increase of ~2%), effects that were repeatable within subjects in a single session (N = 2).
Pinacidil does not have direct effect on nonvascular tissue
Pinacidil does not impact spiking probability or input resistance in neurons or membrane potential in neurons and astrocytes
In recordings from regular-spiking neurons of pyramidal shape (N = 30), we saw no change in any metric measured. At 50 seconds after application, approximately the time of peak vasodilatory effects in vivo, the resting membrane potential did not change (variation of −0.5 ± 0.9 mV standard deviation; Figure 2E), the spike rate induced by current injection did not change (variation of 0.1 ± 1.1 spikes/stimulation; Figure 2F), and input resistance did not change (variation of −0.7 ± 6.5 MΩ; Figure 2D). Similarly, in a limited subset of recordings from fast-spiking interneurons (N = 3), we did not observe any impact of pinacidil application. All significance tests were paired t tests (P > .10).
Astrocytes (N = 35) also showed no significant effects of pinacidil application, demonstrating only a slow depolarization during application of ethanol (1.3 ± 1.7 mV at 50 seconds after drug application) and pinacidil with ethanol (0.5 ± 2.1 mV) (Figure 2B). When we plotted the change in membrane potential at 10-second intervals since drug application (0 to 80 seconds post-drug), we found no trends in astrocytic response to either ACSF or pinacidil (Figure 2C).
During recordings at double our typical application dose (800 μM), we observed 2 pyramidal cells (out of 8) that showed depolarization (peak of 10 to 15 mV) and a loss of spike initiation capability. Following washout, these cells recovered membrane potential but spiking responses to current injection remained impaired. We did not evaluate the impact of this dose on vascular tone or rhythmic vasomotion.
In contrast to the absence of a detectable impact of pinacidil, we found that the membrane potentials of neurons and astrocytes were sensitive to flow rate. Decreasing flow rate caused a consistent depolarization of up to approximately 10 mV that showed an immediate onset, reaching a new baseline within 2 to 5 seconds; increasing flow rate had the opposite effect. In preliminary experiments, we observed two astrocytes that depolarized on switching to the pinacidil solution. These two recordings were obtained prior to placement of an inline pressure meter in the flow pathway that allowed us to monitor and exclude trials that showed flow changes. In the 35 subsequent recordings that did not have flow changes, we never observed a detectable impact of either ethanol or pinacidil on astroctyes or neurons. We also noted that neurons and astrocytes were more likely to die and/or to lose recording quality during a cycle of ethanol or pinacidil presentation, as opposed to ACSF presentation.
DISCUSSION
Pinacidil provides an effective means of inducing vasodilation in vivo. At concentrations less than 400 μM, pinacidil is also selective for cortical vascular smooth muscle, exhibiting no direct effect on intrinsic properties of neurons or astrocytes. As an independent means to induce increased vasodilation and blood volume in a manner analogous to that seen in functional hyperemia, pinacidil provides a viable method for testing the impact of hyperemic events on neural or astrocytic activity. Pinacidil may also be a selective means of emulating other normal hemodynamic phenomena and could have therapeutic applications, such as targeted administration of pinacidil in response to acute vessel obstruction to maintain sufficient perfusion.
The hemodynamic effects induced by pinacidil are similar to natural functional hyperemia. In SI during sensory stimulation in rodents, increases in total oxygenated hemoglobin during sensory stimulation— analogous to our measurement of cortical darkening at 550 nm—peak in a range of 2% to 5%,20–22 and arteries/arterioles dilate 10% to 20%.23 The time course of pinacidil’s effects also parallels the sustained response to continued sensory drive. Arterial diameter in rodent SI and the blood oxygen level–dependent response on functional magnetic resonance imaging in humans and rodents remain high when tactile input is sustained for periods lasting tens of seconds,23,24 as they do under pinacidil application.
Although pinacidil represents an important step forward in our ability to control blood flow while probing the impact of hemodynamics in cortex, it has limitations. The drug is only capable of producing vasodilation; drugs in the same family that block the SUR2B channels to create vasoconstriction (such as diazoxide or glibenclamide) or thromboxane receptor agonists18,25,26 are unfortunately known to be nonspecific, affecting neurons as well as blood vessels. Pinacidil is also not water-soluble, requiring its dissolution in ethanol or DMSO, agents that can have confounding impacts on the system. Applied in vivo, pinacidil also does not appear to wash out fully, or its impact on smooth muscles persists, so that the first trial in each animal is the most consistent and effective one. These limitations stated, this pharmacological approach nevertheless represents a unique means of selective hyperemia induction in vivo.
- Roy CS, Sherrington CS. On the regulation of the blood-supply of the brain. J Physiol 1890; 11:85–108,158-7–158-17.
- Raichle ME. Behind the scenes of functional brain imaging: a historical and physiological perspective. Proc Natl Acad Sci USA 1998; 95:765–772.
- Moore CI, Cao R. The hemo-neural hypothesis: on the role of blood flow in information processing. J Neurophysiol 2008; 99:2035–2047.
- Garthwaite G, Bartus K, Malcolm D, et al. Signaling from blood vessels to CNS axons through nitric oxide. J Neurosci 2006; 26:7730–7740.
- Kozlov AS, Angulo MC, Audinat E, Charpak S. Target cellspecific modulation of neuronal activity by astrocytes. Proc Natl Acad Sci USA 2006; 103:10058–10063.
- Nedergaard M, Ransom B, Goldman SA. New roles for astrocytes: redefining the functional architecture of the brain. Trends Neurosci 2003; 26:523–530.
- Haydon PG, Carmignoto G. Astrocyte control of synaptic transmission and neurovascular coupling. Physiol Rev 2006; 86:1009–1031.
- Ashcroft FM, Gribble FM. New windows on the mechanism of action of K(ATP) channel openers. Trends Pharmacol Sci 2000; 21:439–445.
- Levin BE, Dunn-Meynell AA, Routh VH. Brain glucosensing and the K(ATP) channel. Nat Neurosci 2001; 4:459–460.
- Zawar C, Plant TD, Schirra C, Konnerth A, Neumcke B. Celltype specific expression of ATP-sensitive potassium channels in the rat hippocampus. J Physiol 1999; 514:327–341.
- Li L, Wu J, Jiang C. Differential expression of Kir6.1 and SUR2B mRNAs in the vasculature of various tissues in rats. J Membr Biol 2003; 196:61–69.
- Schwanstecher M, Sieverding C, Dorschner H, et al. Potassium channel openers require ATP to bind to and act through sulfonylurea receptors. EMBO J 1998; 17:5529–5535.
- Shindo T, Yamada M, Isomoto S, Horio Y, Kurachi Y. SUR2 subtype (A and B)-dependent differential activation of the cloned ATP-sensitive K+ channels by pinacidil and nicorandil. Br J Pharmacol 1998; 124:985–991.
- Russ U, Lange U, Loffler-Walz C, Hambrock A, Quast U. Binding and effect of KATP channel openers in the absence of Mg2+. Br J Pharmacol 2003; 139:368–380.
- Higdon NR, Khan SA, Buchanan LV, Meisheri KD. Tissue and species variation in the vascular receptor binding of 3H-P1075, a potent KATP opener vasodilator. J Pharmacol Exp Ther 1997; 280:255–260.
- Wahl M. The effects of pinacidil and tolbutamide in feline pial arteries in situ. Pflugers Arch 1989; 415:250–252.
- Hempelmann RG, Barth HL, Mehdorn HM, Pradel RH, Ziegler A. Effects of potassium channel openers in isolated human cerebral arteries. Neurosurgery 1995; 37:1146–1153.
- Quayle JM, Nelson MT, Standen NB. ATP-sensitive and inwardly rectifying potassium channels in smooth muscle. Physiol Rev 1997; 77:1165–1232.
- Nimmerjahn A, Kirchhoff F, Kerr JN, Helmchen F. Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat Methods 2004; 1:31–37.
- Kong Y, Zheng Y, Johnston D, et al. A model of the dynamic relationship between blood flow and volume changes during brain activation. J Cereb Blood Flow Metab 2004; 24:1382–1392.
- Martin C, Martindale J, Berwick J, Mayhew J. Investigating neural-hemodynamic coupling and the hemodynamic response function in the awake rat. Neuroimage 2006; 32:33–48.
- Jones M, Devonshire IM, Berwick J, Martin C, Redgrave P, Mayhew J. Alterered neurovascular coupling during information-processing states. Eur J Neurosci 2008; 27:2758–2772.
- Woolsey TA, Rovainen CM, Cox SB, et al. Neuronal units linked to microvascular modules in cerebral cortex: response elements for imaging the brain. Cereb Cortex 1996; 6:647–660.
- Moore CI, Stern CE, Corkin S, et al. Segregation of somatosensory activation in the human rolandic cortex using fMRI. J Neurophysiol 2000; 84:558–569.
- Crépel V, Krnjević K, Ben-Ari Y. Sulphonylureas reduce the slowly inactivating D-type outward current in rat hippocampal neurons. J Physiol 1993; 466:39–54.
- Lovick TA, Brown LA, Key BJ. Neuronal activity-related coupling in cortical arterioles: involvement of astrocyte-derived factors. Exp Physiol 2005; 90:131–140.
- Roy CS, Sherrington CS. On the regulation of the blood-supply of the brain. J Physiol 1890; 11:85–108,158-7–158-17.
- Raichle ME. Behind the scenes of functional brain imaging: a historical and physiological perspective. Proc Natl Acad Sci USA 1998; 95:765–772.
- Moore CI, Cao R. The hemo-neural hypothesis: on the role of blood flow in information processing. J Neurophysiol 2008; 99:2035–2047.
- Garthwaite G, Bartus K, Malcolm D, et al. Signaling from blood vessels to CNS axons through nitric oxide. J Neurosci 2006; 26:7730–7740.
- Kozlov AS, Angulo MC, Audinat E, Charpak S. Target cellspecific modulation of neuronal activity by astrocytes. Proc Natl Acad Sci USA 2006; 103:10058–10063.
- Nedergaard M, Ransom B, Goldman SA. New roles for astrocytes: redefining the functional architecture of the brain. Trends Neurosci 2003; 26:523–530.
- Haydon PG, Carmignoto G. Astrocyte control of synaptic transmission and neurovascular coupling. Physiol Rev 2006; 86:1009–1031.
- Ashcroft FM, Gribble FM. New windows on the mechanism of action of K(ATP) channel openers. Trends Pharmacol Sci 2000; 21:439–445.
- Levin BE, Dunn-Meynell AA, Routh VH. Brain glucosensing and the K(ATP) channel. Nat Neurosci 2001; 4:459–460.
- Zawar C, Plant TD, Schirra C, Konnerth A, Neumcke B. Celltype specific expression of ATP-sensitive potassium channels in the rat hippocampus. J Physiol 1999; 514:327–341.
- Li L, Wu J, Jiang C. Differential expression of Kir6.1 and SUR2B mRNAs in the vasculature of various tissues in rats. J Membr Biol 2003; 196:61–69.
- Schwanstecher M, Sieverding C, Dorschner H, et al. Potassium channel openers require ATP to bind to and act through sulfonylurea receptors. EMBO J 1998; 17:5529–5535.
- Shindo T, Yamada M, Isomoto S, Horio Y, Kurachi Y. SUR2 subtype (A and B)-dependent differential activation of the cloned ATP-sensitive K+ channels by pinacidil and nicorandil. Br J Pharmacol 1998; 124:985–991.
- Russ U, Lange U, Loffler-Walz C, Hambrock A, Quast U. Binding and effect of KATP channel openers in the absence of Mg2+. Br J Pharmacol 2003; 139:368–380.
- Higdon NR, Khan SA, Buchanan LV, Meisheri KD. Tissue and species variation in the vascular receptor binding of 3H-P1075, a potent KATP opener vasodilator. J Pharmacol Exp Ther 1997; 280:255–260.
- Wahl M. The effects of pinacidil and tolbutamide in feline pial arteries in situ. Pflugers Arch 1989; 415:250–252.
- Hempelmann RG, Barth HL, Mehdorn HM, Pradel RH, Ziegler A. Effects of potassium channel openers in isolated human cerebral arteries. Neurosurgery 1995; 37:1146–1153.
- Quayle JM, Nelson MT, Standen NB. ATP-sensitive and inwardly rectifying potassium channels in smooth muscle. Physiol Rev 1997; 77:1165–1232.
- Nimmerjahn A, Kirchhoff F, Kerr JN, Helmchen F. Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat Methods 2004; 1:31–37.
- Kong Y, Zheng Y, Johnston D, et al. A model of the dynamic relationship between blood flow and volume changes during brain activation. J Cereb Blood Flow Metab 2004; 24:1382–1392.
- Martin C, Martindale J, Berwick J, Mayhew J. Investigating neural-hemodynamic coupling and the hemodynamic response function in the awake rat. Neuroimage 2006; 32:33–48.
- Jones M, Devonshire IM, Berwick J, Martin C, Redgrave P, Mayhew J. Alterered neurovascular coupling during information-processing states. Eur J Neurosci 2008; 27:2758–2772.
- Woolsey TA, Rovainen CM, Cox SB, et al. Neuronal units linked to microvascular modules in cerebral cortex: response elements for imaging the brain. Cereb Cortex 1996; 6:647–660.
- Moore CI, Stern CE, Corkin S, et al. Segregation of somatosensory activation in the human rolandic cortex using fMRI. J Neurophysiol 2000; 84:558–569.
- Crépel V, Krnjević K, Ben-Ari Y. Sulphonylureas reduce the slowly inactivating D-type outward current in rat hippocampal neurons. J Physiol 1993; 466:39–54.
- Lovick TA, Brown LA, Key BJ. Neuronal activity-related coupling in cortical arterioles: involvement of astrocyte-derived factors. Exp Physiol 2005; 90:131–140.
The polyvagal theory: New insights into adaptive reactions of the autonomic nervous system
HISTORICAL PERSPECTIVES ON THE AUTONOMIC NERVOUS SYSTEM
Central nervous system regulation of visceral organs is the focus of several historic publications that have shaped the texture of physiological inquiry. For example, in 1872 Darwin acknowledged the dynamic neural relationship between the heart and the brain:
Although Darwin acknowledged the bidirectional communication between the viscera and the brain, subsequent formal description of the autonomic nervous system (eg, by Langley2) minimized the importance of central regulatory structures and afferents. Following Langley, medical and physiological research tended to focus on the peripheral motor nerves of the autonomic nervous sytem, with a conceptual emphasis on the paired antagonism between sympathetic and parasympathetic efferent pathways on the target visceral organs. This focus minimized interest in both afferent pathways and the brainstem areas that regulate specific efferent pathways.
The early conceptualization of the vagus focused on an undifferentiated efferent pathway that was assumed to modulate “tone” concurrently to several target organs. Thus, brainstem areas regulating the supradiaphragmatic (eg, myelinated vagal pathways originating in the nucleus ambiguus and terminating primarily above the diaphragm) were not functionally distinguished from those regulating the subdiaphragmatic (eg, unmyelinated vagal pathways originating in the dorsal motor nucleus of the vagus and terminating primarily below the diaphragm). Without this distinction, research and theory focused on the paired antagonism between the parasympathetic and sympathetic innervation to target organs. The consequence of an emphasis on paired antagonism was an acceptance in physiology and medicine of global constructs such as autonomic balance, sympathetic tone, and vagal tone.
More than 50 years ago, Hess proposed that the autonomic nervous system was not solely vegetative and automatic but was instead an integrated system with both peripheral and central neurons.3 By emphasizing the central mechanisms that mediate the dynamic regulation of peripheral organs, Hess anticipated the need for technologies to continuously monitor peripheral and central neural circuits involved in the regulation of visceral function.
THE VAGAL PARADOX
In 1992, I proposed that an estimate of vagal tone, derived from measuring respiratory sinus arrhythmia, could be used in clinical medicine as an index of stress vulnerability.4 Rather than using the descriptive measures of heart rate variability (ie, beat-to-beat variability) frequently used in obstetrics and pediatrics, the paper emphasized that respiratory sinus arrhythmia has a neural origin and represents the tonic functional outflow from the vagus to the heart (ie, cardiac vagal tone). Thus, it was proposed that respiratory sinus arrhythmia would provide a more sensitive index of health status than a more global measure of beat-to-beat heart rate variability reflecting undetermined neural and nonneural mechanisms. The paper presented a quantitative approach that applied time-series analyses to extract the amplitude of respiratory sinus arrhythmia as a more accurate index of vagal activity. The article provided data demonstrating that healthy full-term infants had respiratory sinus arrhythmia of significantly greater amplitude than did preterm infants. This idea of using heart rate patterns to index vagal activity was not new, having been reported as early as 1910 by Hering.5 Moreover, contemporary studies have reliably reported that vagal blockade via atropine depresses respiratory sinus arrhythmia in mammals.6,7
In response to this article,4 I received a letter from a neonatologist who wrote that, as a medical student, he learned that vagal tone could be lethal. He argued that perhaps too much of a good thing (ie, vagal tone) could be bad. He was referring, of course, to the clinical risk of neurogenic bradycardia. Bradycardia, when observed during delivery, may be an indicator of fetal distress. Similarly, bradycardia and apnea are important indicators of risk for the newborn.
My colleagues and I further investigated this perplexing observation by studying the human fetus during delivery. We observed that fetal bradycardia occurred only when respiratory sinus arrhythmia was depressed (ie, a respiratory rhythm in fetal heart rate is observable even in the absence of the large chest wall movements associated with breathing that occur postpartum).8 This raised the question of how vagal mechanisms could mediate both respiratory sinus arrhythmia and bradycardia, as one is protective and the other is potentially lethal. This inconsistency became the “vagal paradox” and served as the motivation behind the polyvagal theory.
With regard to the mechanisms mediating bradycardia and heart rate variability, there is an obvious inconsistency between data and physiological assumptions. Physiological models assume vagal regulation of both chronotropic control of the heart (ie, heart rate) and the amplitude of respiratory sinus arrhythmia.9,10 For example, it has been reliably reported that vagal cardio-inhibitory fibers to the heart have consistent functional properties characterized by bradycardia to neural stimulation and a respiratory rhythm.9 However, although there are situations in which both measures covary (eg, during exercise and cholinergic blockade), there are other situations in which the measures appear to reflect independent sources of neural control (eg, bradycardic episodes associated with hypoxia, vasovagal syncope, and fetal distress). In contrast to these observable phenomena, researchers continue to argue for a covariation between these two parameters. This inconsistency, based on an assumption of a single central vagal source, is what I have labeled the vagal paradox.
THE POLYVAGAL THEORY: THREE PHYLOGENETIC RESPONSE SYSTEMS
Investigation of the phylogeny of the vertebrate autonomic nervous system provides an answer to the vagal paradox. Research in comparative neuroanatomy and neurophysiology has identified two branches of the vagus, with each branch supporting different adaptive functions and behavioral strategies. The vagal output to the heart from one branch is manifested in respiratory sinus arrhythmia, and the output from the other branch is manifested in bradycardia and possibly the slower rhythms in heart rate variability. Although the slower rhythms have been assumed to have a sympathetic influence, they are blocked by atropine.7
The polyvagal theory7,11–15 articulates how each of three phylogenetic stages in the development of the vertebrate autonomic nervous system is associated with a distinct autonomic subsystem that is retained and expressed in mammals. These autonomic subsystems are phylogenetically ordered and behaviorally linked to social communication (eg, facial expression, vocalization, listening), mobilization (eg, fight–flight behaviors), and immobilization (eg, feigning death, vasovagal syncope, and behavioral shutdown).
The social communication system (ie, social engagement system; see below) involves the myelinated vagus, which serves to foster calm behavioral states by inhibiting sympathetic influences to the heart and dampening the hypothalamic-pituitary-adrenal (HPA) axis.16 The mobilization system is dependent on the functioning of the sympathetic nervous system. The most phylogenetically primitive component, the immobilization system, is dependent on the unmyelinated vagus, which is shared with most vertebrates. With increased neural complexity resulting from phylogenetic development, the organism’s behavioral and affective repertoire is enriched. The three circuits can be conceptualized as dynamic, providing adaptive responses to safe, dangerous, and life-threatening events and contexts.
Only mammals have a myelinated vagus. Unlike the unmyelinated vagus, originating in the dorsal motor nucleus of the vagus with pre- and postganglionic muscarinic receptors, the mammalian myelinated vagus originates in the nucleus ambiguus and has preganglionic nicotinic receptors and postganglionic muscarinic receptors. The unmyelinated vagus is shared with other vertebrates, including reptiles, amphibians, teleosts, and elasmobranchs.
We are now investigating the possibility of extracting different features of the heart rate pattern to dynamically monitor the two vagal systems. Preliminary studies in our laboratory support this possibility. In these studies we have blocked the nicotinic preganglionic receptors with hexamethonium and the muscarinic receptors with atropine. The data were collected from the prairie vole,17 which has a very high ambient vagal tone. These preliminary data demonstrated that, in several animals, nicotinic blockade selectively removes respiratory sinus arrhythmia without dampening the amplitude of the lower frequencies in heart rate variability. In contrast, blocking the muscarinic receptors with atropine removes both the low and respiratory frequencies.
CONSISTENCY WITH JACKSONIAN DISSOLUTION
The three circuits are organized and respond to challenges in a phylogenetically determined hierarchy consistent with the Jacksonian principle of dissolution. Jackson proposed that in the brain, higher (ie, phylogenetically newer) neural circuits inhibit lower (ie, phylogenetically older) neural circuits and “when the higher are suddenly rendered functionless, the lower rise in activity.” 18 Although Jackson proposed dissolution to explain changes in brain function due to damage and illness, the polyvagal theory proposes a similar phylogenetically ordered hierarchical model to describe the sequence of autonomic response strategies to challenges.
The human nervous system, similar to that of other mammals, evolved not solely to survive in safe environments but also to promote survival in dangerous and life-threatening contexts. To accomplish this adaptive flexibility, the human nervous system retained two more primitive neural circuits to regulate defensive strategies (ie, fight–flight and death-feigning behaviors). It is important to note that social behavior, social communication, and visceral homeostasis are incompatible with the neurophysiological states and behaviors promoted by the two neural circuits that support defense strategies. Thus, via evolution, the human nervous system retains three neural circuits, which are in a phylogenetically organized hierarchy. In this hierarchy of adaptive responses, the newest circuit is used first; if that circuit fails to provide safety, the older circuits are recruited sequentially.
Investigation of the phylogeny of regulation of the vertebrate heart11,12,19,20 has led to extraction of four principles that provide a basis for testing of hypotheses relating specific neural mechanisms to social engagement, fight–flight, and death-feigning behaviors:
- There is a phylogenetic shift in the regulation of the heart from endocrine communication to unmyelinated nerves and finally to myelinated nerves.
- There is a development of opposing neural mechanisms of excitation and inhibition to provide rapid regulation of graded metabolic output.
- A face–heart connection evolved as source nuclei of vagal pathways shifted ventrally from the older dorsal motor nucleus to the nucleus ambiguus. This resulted in an anatomical and neurophysiological linkage between neural regulation of the heart via the myelinated vagus and the special visceral efferent pathways that regulate the striated muscles of the face and head, forming an integrated social engagement system (Figure 1; for more details, see Porges7,15).
- With increased cortical development, the cortex exhibits greater control over the brainstem via direct (eg, corticobulbar) and indirect (eg, corticoreticular) neural pathways originating in motor cortex and terminating in the source nuclei of the myelinated motor nerves emerging from the brainstem (eg, specific neural pathways embedded within cranial nerves V, VII, IX, X, and XI), controlling visceromotor structures (ie, heart, bronchi) as well as somatomotor structures (muscles of the face and head).
NEUROCEPTION: CONTEXTUAL CUEING OF ADAPTIVE, MALADAPTIVE PHYSIOLOGICAL STATES
To effectively switch from defensive to social engagement strategies, the mammalian nervous system needs to perform two important adaptive tasks: (1) assess risk, and (2) if the environment is perceived as safe, inhibit the more primitive limbic structures that control fight, flight, or freeze behaviors.
Any stimulus that has the potential for increasing an organism’s experience of safety has the potential of recruiting the evolutionarily more advanced neural circuits that support the prosocial behaviors of the social engagement system.
The nervous system, through the processing of sensory information from the environment and from the viscera, continuously evaluates risk. Since the neural evaluation of risk does not require conscious awareness and may involve subcortical limbic structures,21 the term neuroception22 was introduced to emphasize a neural process, distinct from perception, that is capable of distinguishing environmental (and visceral) features that are safe, dangerous, or life-threatening. In safe environments, autonomic state is adaptively regulated to dampen sympathetic activation and to protect the oxygen-dependent central nervous system, especially the cortex, from the metabolically conservative reactions of the dorsal vagal complex. However, how does the nervous system know when the environment is safe, dangerous, or life-threatening, and which neural mechanisms evaluate this risk?
Environmental components of neuroception
Neuroception represents a neural process that enables humans and other mammals to engage in social behaviors by distinguishing safe from dangerous contexts. Neuroception is proposed as a plausible mechanism mediating both the expression and the disruption of positive social behavior, emotion regulation, and visceral homeostasis.7,22 Neuroception might be triggered by feature detectors involving areas of temporal cortex that communicate with the central nucleus of the amygdala and the periaqueductal gray, since limbic reactivity is modulated by temporal cortex responses to the intention of voices, faces, and hand movements. Thus, the neuroception of familiar individuals and individuals with appropriately prosodic voices and warm, expressive faces translates into a social interaction promoting a sense of safety.
In most individuals (ie, those without a psychiatric disorder or neuropathology), the nervous system evaluates risk and matches neurophysiological state with the actual risk of the environment. When the environment is appraised as being safe, the defensive limbic structures are inhibited, enabling social engagement and calm visceral states to emerge. In contrast, some individuals experience a mismatch and the nervous system appraises the environment as being dangerous even when it is safe. This mismatch results in physiological states that support fight, flight, or freeze behaviors, but not social engagement behaviors. According to the theory, social communication can be expressed efficiently through the social engagement system only when these defensive circuits are inhibited.
Other contributors to neuroception
The features of risk in the environment do not solely drive neuroception. Afferent feedback from the viscera provides a major mediator of the accessibility of prosocial circuits associated with social engagement behaviors. For example, the polyvagal theory predicts that states of mobilization would compromise our ability to detect positive social cues. Functionally, visceral states color our perception of objects and others. Thus, the same features of one person engaging another may result in a range of outcomes, depending on the physiological state of the target individual. If the person being engaged is in a state in which the social engagement system is easily accessible, the reciprocal prosocial interactions are likely to occur. However, if the individual is in a state of mobilization, the same engaging response might be responded to with the asocial features of withdrawal or aggression. In such a state, it might be very difficult to dampen the mobilization circuit and enable the social engagement system to come back on line.
The insula may be involved in the mediation of neuroception, since it has been proposed as a brain structure involved in conveying the diffuse feedback from the viscera into cognitive awareness. Functional imaging experiments have demonstrated that the insula plays an important role in the experience of pain and the experience of several emotions, including anger, fear, disgust, happiness, and sadness. Critchley proposes that internal body states are represented in the insula and contribute to states of subjective feeling, and he has demonstrated that activity in the insula correlates with interoceptive accuracy.23
SUMMARY
The polyvagal theory proposes that the evolution of the mammalian autonomic nervous system provides the neurophysiological substrates for adaptive behavioral strategies. It further proposes that physiological state limits the range of behavior and psychological experience. The theory links the evolution of the autonomic nervous system to affective experience, emotional expression, facial gestures, vocal communication, and contingent social behavior. In this way, the theory provides a plausible explanation for the reported covariation between atypical autonomic regulation (eg, reduced vagal and increased sympathetic influences to the heart) and psychiatric and behavioral disorders that involve difficulties in regulating appropriate social, emotional, and communication behaviors.
The polyvagal theory provides several insights into the adaptive nature of physiological state. First, the theory emphasizes that physiological states support different classes of behavior. For example, a physiological state characterized by a vagal withdrawal would support the mobilization behaviors of fight and flight. In contrast, a physiological state characterized by increased vagal influence on the heart (via myelinated vagal pathways originating in the nucleus ambiguus) would support spontaneous social engagement behaviors. Second, the theory emphasizes the formation of an integrated social engagement system through functional and structural links between neural control of the striated muscles of the face and the smooth muscles of the viscera. Third, the polyvagal theory proposes a mechanism—neuroception— to trigger or to inhibit defense strategies.
- Darwin C. The Expression of Emotions in Man and Animals. New York, NY: D Appleton;1872.
- Langley JN. The Autonomic Nervous System. Cambridge, England: Heffer & Sons; 1921.
- Hess WR. Diencephalon, Autonomic and Extrapyramidal Functions. New York, NY: Grune & Stratton; 1954.
- Porges SW. Vagal tone: a physiologic marker of stress vulnerability. Pediatrics 1992; 90:498–504.
- Hering HE. A functional test of heart vagi in man. Menschen Munchen Medizinische Wochenschrift 1910; 57:1931–1933.
- Porges SW. Respiratory sinus arrhythmia: physiological basis, quantitative methods, and clinical implications. In:Grossman P, Janssen K, Vaitl D, eds. Cardiorespiratory and Cardiosomatic Psychophysiology. New York, NY: Plenum; 1986:101–115.
- Porges SW. The polyvagal perspective. Biol Psychol 2007; 74:116–143.
- Reed SF, Ohel G, David R, Porges SW. A neural explanation of fetal heart rate patterns: a test of the polyvagal theory. Dev Psychobiol 1999; 35:108–118.
- Jordan D, Khalid ME, Schneiderman N, Spyer KM. The location and properties of preganglionic vagal cardiomotor neurones in the rabbit. Pflugers Arch 1982; 395:244–250.
- Katona PG, Jih F. Respiratory sinus arrhythmia: noninvasive measure of parasympathetic cardiac control. J Appl Physiol 1975; 39:801–805.
- Porges SW. Orienting in a defensive world: mammalian modifications of our evolutionary heritage—a polyvagal theory. Psychophysiology 1995; 32:301–318.
- Porges SW. Emotion: An evolutionary by-product of the neural regulation of the autonomic nervous system. Ann N Y Acad Sci 1997; 807:62–77.
- Porges SW. Love: An emergent property of the mammalian autonomic nervous system. Psychoneuroendocrinology 1998; 23:837–861.
- Porges SW. The polyvagal theory: phylogenetic substrates of a social nervous system. Int J Psychophysiol 2001; 42:123–146.
- Porges SW. Social engagement and attachment: a phylogenetic perspective. Ann N Y Acad Sci 2003; 1008:31–47.
- Bueno L, Gue M, Fargeas MJ, Alvinerie M, Junien JL, Fioramonti J. Vagally mediated inhibition of acoustic stress-induced cortisol release by orally administered kappa-opioid substances in dogs. Endocrinology 1989; 124:1788–1793.
- Grippo AJ, Lamb DG, Carter CS, Porges SW. Cardiac regulation in the socially monogamous prairie vole. Physiol Behav 2007; 90:386–393.
- Jackson JH. Evolution and dissolution of the nervous system. In:Taylor J, ed. Selected Writings of John Hughlings Jackson. London: Stapes Press; 1958:45–118.
- Morris JL, Nilsson S. The circulatory system. In:Nilsson S, Holmgren S, eds. Comparative Physiology and Evolution of the Autonomic Nervous System. Chur, Switzerland: Harwood Academic Publishers; 1994.
- Taylor EW, Jordan D, Coote JH. Central control of the cardiovascular and respiratory systems and their interactions in vertebrates. Physiol Rev 1999; 79:855–916.
- Morris JS, Ohman A, Dolan RJ. A subcortical pathway to the right amygdala mediating “unseen” fear. Proc Natl Acad Sci U S A 1999; 96:1680–1685.
- Porges SW. Neuroception: a subconscious system for detecting threat and safety. Zero to Three: Bulletin of the National Center for Clinical Infant Programs 2004; 24( 5):19–24.
- Critchley HD. Neural mechanisms of autonomic, affective, and cognitive integration. J Comp Neurol 2005; 493:154–166.
HISTORICAL PERSPECTIVES ON THE AUTONOMIC NERVOUS SYSTEM
Central nervous system regulation of visceral organs is the focus of several historic publications that have shaped the texture of physiological inquiry. For example, in 1872 Darwin acknowledged the dynamic neural relationship between the heart and the brain:
Although Darwin acknowledged the bidirectional communication between the viscera and the brain, subsequent formal description of the autonomic nervous system (eg, by Langley2) minimized the importance of central regulatory structures and afferents. Following Langley, medical and physiological research tended to focus on the peripheral motor nerves of the autonomic nervous sytem, with a conceptual emphasis on the paired antagonism between sympathetic and parasympathetic efferent pathways on the target visceral organs. This focus minimized interest in both afferent pathways and the brainstem areas that regulate specific efferent pathways.
The early conceptualization of the vagus focused on an undifferentiated efferent pathway that was assumed to modulate “tone” concurrently to several target organs. Thus, brainstem areas regulating the supradiaphragmatic (eg, myelinated vagal pathways originating in the nucleus ambiguus and terminating primarily above the diaphragm) were not functionally distinguished from those regulating the subdiaphragmatic (eg, unmyelinated vagal pathways originating in the dorsal motor nucleus of the vagus and terminating primarily below the diaphragm). Without this distinction, research and theory focused on the paired antagonism between the parasympathetic and sympathetic innervation to target organs. The consequence of an emphasis on paired antagonism was an acceptance in physiology and medicine of global constructs such as autonomic balance, sympathetic tone, and vagal tone.
More than 50 years ago, Hess proposed that the autonomic nervous system was not solely vegetative and automatic but was instead an integrated system with both peripheral and central neurons.3 By emphasizing the central mechanisms that mediate the dynamic regulation of peripheral organs, Hess anticipated the need for technologies to continuously monitor peripheral and central neural circuits involved in the regulation of visceral function.
THE VAGAL PARADOX
In 1992, I proposed that an estimate of vagal tone, derived from measuring respiratory sinus arrhythmia, could be used in clinical medicine as an index of stress vulnerability.4 Rather than using the descriptive measures of heart rate variability (ie, beat-to-beat variability) frequently used in obstetrics and pediatrics, the paper emphasized that respiratory sinus arrhythmia has a neural origin and represents the tonic functional outflow from the vagus to the heart (ie, cardiac vagal tone). Thus, it was proposed that respiratory sinus arrhythmia would provide a more sensitive index of health status than a more global measure of beat-to-beat heart rate variability reflecting undetermined neural and nonneural mechanisms. The paper presented a quantitative approach that applied time-series analyses to extract the amplitude of respiratory sinus arrhythmia as a more accurate index of vagal activity. The article provided data demonstrating that healthy full-term infants had respiratory sinus arrhythmia of significantly greater amplitude than did preterm infants. This idea of using heart rate patterns to index vagal activity was not new, having been reported as early as 1910 by Hering.5 Moreover, contemporary studies have reliably reported that vagal blockade via atropine depresses respiratory sinus arrhythmia in mammals.6,7
In response to this article,4 I received a letter from a neonatologist who wrote that, as a medical student, he learned that vagal tone could be lethal. He argued that perhaps too much of a good thing (ie, vagal tone) could be bad. He was referring, of course, to the clinical risk of neurogenic bradycardia. Bradycardia, when observed during delivery, may be an indicator of fetal distress. Similarly, bradycardia and apnea are important indicators of risk for the newborn.
My colleagues and I further investigated this perplexing observation by studying the human fetus during delivery. We observed that fetal bradycardia occurred only when respiratory sinus arrhythmia was depressed (ie, a respiratory rhythm in fetal heart rate is observable even in the absence of the large chest wall movements associated with breathing that occur postpartum).8 This raised the question of how vagal mechanisms could mediate both respiratory sinus arrhythmia and bradycardia, as one is protective and the other is potentially lethal. This inconsistency became the “vagal paradox” and served as the motivation behind the polyvagal theory.
With regard to the mechanisms mediating bradycardia and heart rate variability, there is an obvious inconsistency between data and physiological assumptions. Physiological models assume vagal regulation of both chronotropic control of the heart (ie, heart rate) and the amplitude of respiratory sinus arrhythmia.9,10 For example, it has been reliably reported that vagal cardio-inhibitory fibers to the heart have consistent functional properties characterized by bradycardia to neural stimulation and a respiratory rhythm.9 However, although there are situations in which both measures covary (eg, during exercise and cholinergic blockade), there are other situations in which the measures appear to reflect independent sources of neural control (eg, bradycardic episodes associated with hypoxia, vasovagal syncope, and fetal distress). In contrast to these observable phenomena, researchers continue to argue for a covariation between these two parameters. This inconsistency, based on an assumption of a single central vagal source, is what I have labeled the vagal paradox.
THE POLYVAGAL THEORY: THREE PHYLOGENETIC RESPONSE SYSTEMS
Investigation of the phylogeny of the vertebrate autonomic nervous system provides an answer to the vagal paradox. Research in comparative neuroanatomy and neurophysiology has identified two branches of the vagus, with each branch supporting different adaptive functions and behavioral strategies. The vagal output to the heart from one branch is manifested in respiratory sinus arrhythmia, and the output from the other branch is manifested in bradycardia and possibly the slower rhythms in heart rate variability. Although the slower rhythms have been assumed to have a sympathetic influence, they are blocked by atropine.7
The polyvagal theory7,11–15 articulates how each of three phylogenetic stages in the development of the vertebrate autonomic nervous system is associated with a distinct autonomic subsystem that is retained and expressed in mammals. These autonomic subsystems are phylogenetically ordered and behaviorally linked to social communication (eg, facial expression, vocalization, listening), mobilization (eg, fight–flight behaviors), and immobilization (eg, feigning death, vasovagal syncope, and behavioral shutdown).
The social communication system (ie, social engagement system; see below) involves the myelinated vagus, which serves to foster calm behavioral states by inhibiting sympathetic influences to the heart and dampening the hypothalamic-pituitary-adrenal (HPA) axis.16 The mobilization system is dependent on the functioning of the sympathetic nervous system. The most phylogenetically primitive component, the immobilization system, is dependent on the unmyelinated vagus, which is shared with most vertebrates. With increased neural complexity resulting from phylogenetic development, the organism’s behavioral and affective repertoire is enriched. The three circuits can be conceptualized as dynamic, providing adaptive responses to safe, dangerous, and life-threatening events and contexts.
Only mammals have a myelinated vagus. Unlike the unmyelinated vagus, originating in the dorsal motor nucleus of the vagus with pre- and postganglionic muscarinic receptors, the mammalian myelinated vagus originates in the nucleus ambiguus and has preganglionic nicotinic receptors and postganglionic muscarinic receptors. The unmyelinated vagus is shared with other vertebrates, including reptiles, amphibians, teleosts, and elasmobranchs.
We are now investigating the possibility of extracting different features of the heart rate pattern to dynamically monitor the two vagal systems. Preliminary studies in our laboratory support this possibility. In these studies we have blocked the nicotinic preganglionic receptors with hexamethonium and the muscarinic receptors with atropine. The data were collected from the prairie vole,17 which has a very high ambient vagal tone. These preliminary data demonstrated that, in several animals, nicotinic blockade selectively removes respiratory sinus arrhythmia without dampening the amplitude of the lower frequencies in heart rate variability. In contrast, blocking the muscarinic receptors with atropine removes both the low and respiratory frequencies.
CONSISTENCY WITH JACKSONIAN DISSOLUTION
The three circuits are organized and respond to challenges in a phylogenetically determined hierarchy consistent with the Jacksonian principle of dissolution. Jackson proposed that in the brain, higher (ie, phylogenetically newer) neural circuits inhibit lower (ie, phylogenetically older) neural circuits and “when the higher are suddenly rendered functionless, the lower rise in activity.” 18 Although Jackson proposed dissolution to explain changes in brain function due to damage and illness, the polyvagal theory proposes a similar phylogenetically ordered hierarchical model to describe the sequence of autonomic response strategies to challenges.
The human nervous system, similar to that of other mammals, evolved not solely to survive in safe environments but also to promote survival in dangerous and life-threatening contexts. To accomplish this adaptive flexibility, the human nervous system retained two more primitive neural circuits to regulate defensive strategies (ie, fight–flight and death-feigning behaviors). It is important to note that social behavior, social communication, and visceral homeostasis are incompatible with the neurophysiological states and behaviors promoted by the two neural circuits that support defense strategies. Thus, via evolution, the human nervous system retains three neural circuits, which are in a phylogenetically organized hierarchy. In this hierarchy of adaptive responses, the newest circuit is used first; if that circuit fails to provide safety, the older circuits are recruited sequentially.
Investigation of the phylogeny of regulation of the vertebrate heart11,12,19,20 has led to extraction of four principles that provide a basis for testing of hypotheses relating specific neural mechanisms to social engagement, fight–flight, and death-feigning behaviors:
- There is a phylogenetic shift in the regulation of the heart from endocrine communication to unmyelinated nerves and finally to myelinated nerves.
- There is a development of opposing neural mechanisms of excitation and inhibition to provide rapid regulation of graded metabolic output.
- A face–heart connection evolved as source nuclei of vagal pathways shifted ventrally from the older dorsal motor nucleus to the nucleus ambiguus. This resulted in an anatomical and neurophysiological linkage between neural regulation of the heart via the myelinated vagus and the special visceral efferent pathways that regulate the striated muscles of the face and head, forming an integrated social engagement system (Figure 1; for more details, see Porges7,15).
- With increased cortical development, the cortex exhibits greater control over the brainstem via direct (eg, corticobulbar) and indirect (eg, corticoreticular) neural pathways originating in motor cortex and terminating in the source nuclei of the myelinated motor nerves emerging from the brainstem (eg, specific neural pathways embedded within cranial nerves V, VII, IX, X, and XI), controlling visceromotor structures (ie, heart, bronchi) as well as somatomotor structures (muscles of the face and head).
NEUROCEPTION: CONTEXTUAL CUEING OF ADAPTIVE, MALADAPTIVE PHYSIOLOGICAL STATES
To effectively switch from defensive to social engagement strategies, the mammalian nervous system needs to perform two important adaptive tasks: (1) assess risk, and (2) if the environment is perceived as safe, inhibit the more primitive limbic structures that control fight, flight, or freeze behaviors.
Any stimulus that has the potential for increasing an organism’s experience of safety has the potential of recruiting the evolutionarily more advanced neural circuits that support the prosocial behaviors of the social engagement system.
The nervous system, through the processing of sensory information from the environment and from the viscera, continuously evaluates risk. Since the neural evaluation of risk does not require conscious awareness and may involve subcortical limbic structures,21 the term neuroception22 was introduced to emphasize a neural process, distinct from perception, that is capable of distinguishing environmental (and visceral) features that are safe, dangerous, or life-threatening. In safe environments, autonomic state is adaptively regulated to dampen sympathetic activation and to protect the oxygen-dependent central nervous system, especially the cortex, from the metabolically conservative reactions of the dorsal vagal complex. However, how does the nervous system know when the environment is safe, dangerous, or life-threatening, and which neural mechanisms evaluate this risk?
Environmental components of neuroception
Neuroception represents a neural process that enables humans and other mammals to engage in social behaviors by distinguishing safe from dangerous contexts. Neuroception is proposed as a plausible mechanism mediating both the expression and the disruption of positive social behavior, emotion regulation, and visceral homeostasis.7,22 Neuroception might be triggered by feature detectors involving areas of temporal cortex that communicate with the central nucleus of the amygdala and the periaqueductal gray, since limbic reactivity is modulated by temporal cortex responses to the intention of voices, faces, and hand movements. Thus, the neuroception of familiar individuals and individuals with appropriately prosodic voices and warm, expressive faces translates into a social interaction promoting a sense of safety.
In most individuals (ie, those without a psychiatric disorder or neuropathology), the nervous system evaluates risk and matches neurophysiological state with the actual risk of the environment. When the environment is appraised as being safe, the defensive limbic structures are inhibited, enabling social engagement and calm visceral states to emerge. In contrast, some individuals experience a mismatch and the nervous system appraises the environment as being dangerous even when it is safe. This mismatch results in physiological states that support fight, flight, or freeze behaviors, but not social engagement behaviors. According to the theory, social communication can be expressed efficiently through the social engagement system only when these defensive circuits are inhibited.
Other contributors to neuroception
The features of risk in the environment do not solely drive neuroception. Afferent feedback from the viscera provides a major mediator of the accessibility of prosocial circuits associated with social engagement behaviors. For example, the polyvagal theory predicts that states of mobilization would compromise our ability to detect positive social cues. Functionally, visceral states color our perception of objects and others. Thus, the same features of one person engaging another may result in a range of outcomes, depending on the physiological state of the target individual. If the person being engaged is in a state in which the social engagement system is easily accessible, the reciprocal prosocial interactions are likely to occur. However, if the individual is in a state of mobilization, the same engaging response might be responded to with the asocial features of withdrawal or aggression. In such a state, it might be very difficult to dampen the mobilization circuit and enable the social engagement system to come back on line.
The insula may be involved in the mediation of neuroception, since it has been proposed as a brain structure involved in conveying the diffuse feedback from the viscera into cognitive awareness. Functional imaging experiments have demonstrated that the insula plays an important role in the experience of pain and the experience of several emotions, including anger, fear, disgust, happiness, and sadness. Critchley proposes that internal body states are represented in the insula and contribute to states of subjective feeling, and he has demonstrated that activity in the insula correlates with interoceptive accuracy.23
SUMMARY
The polyvagal theory proposes that the evolution of the mammalian autonomic nervous system provides the neurophysiological substrates for adaptive behavioral strategies. It further proposes that physiological state limits the range of behavior and psychological experience. The theory links the evolution of the autonomic nervous system to affective experience, emotional expression, facial gestures, vocal communication, and contingent social behavior. In this way, the theory provides a plausible explanation for the reported covariation between atypical autonomic regulation (eg, reduced vagal and increased sympathetic influences to the heart) and psychiatric and behavioral disorders that involve difficulties in regulating appropriate social, emotional, and communication behaviors.
The polyvagal theory provides several insights into the adaptive nature of physiological state. First, the theory emphasizes that physiological states support different classes of behavior. For example, a physiological state characterized by a vagal withdrawal would support the mobilization behaviors of fight and flight. In contrast, a physiological state characterized by increased vagal influence on the heart (via myelinated vagal pathways originating in the nucleus ambiguus) would support spontaneous social engagement behaviors. Second, the theory emphasizes the formation of an integrated social engagement system through functional and structural links between neural control of the striated muscles of the face and the smooth muscles of the viscera. Third, the polyvagal theory proposes a mechanism—neuroception— to trigger or to inhibit defense strategies.
HISTORICAL PERSPECTIVES ON THE AUTONOMIC NERVOUS SYSTEM
Central nervous system regulation of visceral organs is the focus of several historic publications that have shaped the texture of physiological inquiry. For example, in 1872 Darwin acknowledged the dynamic neural relationship between the heart and the brain:
Although Darwin acknowledged the bidirectional communication between the viscera and the brain, subsequent formal description of the autonomic nervous system (eg, by Langley2) minimized the importance of central regulatory structures and afferents. Following Langley, medical and physiological research tended to focus on the peripheral motor nerves of the autonomic nervous sytem, with a conceptual emphasis on the paired antagonism between sympathetic and parasympathetic efferent pathways on the target visceral organs. This focus minimized interest in both afferent pathways and the brainstem areas that regulate specific efferent pathways.
The early conceptualization of the vagus focused on an undifferentiated efferent pathway that was assumed to modulate “tone” concurrently to several target organs. Thus, brainstem areas regulating the supradiaphragmatic (eg, myelinated vagal pathways originating in the nucleus ambiguus and terminating primarily above the diaphragm) were not functionally distinguished from those regulating the subdiaphragmatic (eg, unmyelinated vagal pathways originating in the dorsal motor nucleus of the vagus and terminating primarily below the diaphragm). Without this distinction, research and theory focused on the paired antagonism between the parasympathetic and sympathetic innervation to target organs. The consequence of an emphasis on paired antagonism was an acceptance in physiology and medicine of global constructs such as autonomic balance, sympathetic tone, and vagal tone.
More than 50 years ago, Hess proposed that the autonomic nervous system was not solely vegetative and automatic but was instead an integrated system with both peripheral and central neurons.3 By emphasizing the central mechanisms that mediate the dynamic regulation of peripheral organs, Hess anticipated the need for technologies to continuously monitor peripheral and central neural circuits involved in the regulation of visceral function.
THE VAGAL PARADOX
In 1992, I proposed that an estimate of vagal tone, derived from measuring respiratory sinus arrhythmia, could be used in clinical medicine as an index of stress vulnerability.4 Rather than using the descriptive measures of heart rate variability (ie, beat-to-beat variability) frequently used in obstetrics and pediatrics, the paper emphasized that respiratory sinus arrhythmia has a neural origin and represents the tonic functional outflow from the vagus to the heart (ie, cardiac vagal tone). Thus, it was proposed that respiratory sinus arrhythmia would provide a more sensitive index of health status than a more global measure of beat-to-beat heart rate variability reflecting undetermined neural and nonneural mechanisms. The paper presented a quantitative approach that applied time-series analyses to extract the amplitude of respiratory sinus arrhythmia as a more accurate index of vagal activity. The article provided data demonstrating that healthy full-term infants had respiratory sinus arrhythmia of significantly greater amplitude than did preterm infants. This idea of using heart rate patterns to index vagal activity was not new, having been reported as early as 1910 by Hering.5 Moreover, contemporary studies have reliably reported that vagal blockade via atropine depresses respiratory sinus arrhythmia in mammals.6,7
In response to this article,4 I received a letter from a neonatologist who wrote that, as a medical student, he learned that vagal tone could be lethal. He argued that perhaps too much of a good thing (ie, vagal tone) could be bad. He was referring, of course, to the clinical risk of neurogenic bradycardia. Bradycardia, when observed during delivery, may be an indicator of fetal distress. Similarly, bradycardia and apnea are important indicators of risk for the newborn.
My colleagues and I further investigated this perplexing observation by studying the human fetus during delivery. We observed that fetal bradycardia occurred only when respiratory sinus arrhythmia was depressed (ie, a respiratory rhythm in fetal heart rate is observable even in the absence of the large chest wall movements associated with breathing that occur postpartum).8 This raised the question of how vagal mechanisms could mediate both respiratory sinus arrhythmia and bradycardia, as one is protective and the other is potentially lethal. This inconsistency became the “vagal paradox” and served as the motivation behind the polyvagal theory.
With regard to the mechanisms mediating bradycardia and heart rate variability, there is an obvious inconsistency between data and physiological assumptions. Physiological models assume vagal regulation of both chronotropic control of the heart (ie, heart rate) and the amplitude of respiratory sinus arrhythmia.9,10 For example, it has been reliably reported that vagal cardio-inhibitory fibers to the heart have consistent functional properties characterized by bradycardia to neural stimulation and a respiratory rhythm.9 However, although there are situations in which both measures covary (eg, during exercise and cholinergic blockade), there are other situations in which the measures appear to reflect independent sources of neural control (eg, bradycardic episodes associated with hypoxia, vasovagal syncope, and fetal distress). In contrast to these observable phenomena, researchers continue to argue for a covariation between these two parameters. This inconsistency, based on an assumption of a single central vagal source, is what I have labeled the vagal paradox.
THE POLYVAGAL THEORY: THREE PHYLOGENETIC RESPONSE SYSTEMS
Investigation of the phylogeny of the vertebrate autonomic nervous system provides an answer to the vagal paradox. Research in comparative neuroanatomy and neurophysiology has identified two branches of the vagus, with each branch supporting different adaptive functions and behavioral strategies. The vagal output to the heart from one branch is manifested in respiratory sinus arrhythmia, and the output from the other branch is manifested in bradycardia and possibly the slower rhythms in heart rate variability. Although the slower rhythms have been assumed to have a sympathetic influence, they are blocked by atropine.7
The polyvagal theory7,11–15 articulates how each of three phylogenetic stages in the development of the vertebrate autonomic nervous system is associated with a distinct autonomic subsystem that is retained and expressed in mammals. These autonomic subsystems are phylogenetically ordered and behaviorally linked to social communication (eg, facial expression, vocalization, listening), mobilization (eg, fight–flight behaviors), and immobilization (eg, feigning death, vasovagal syncope, and behavioral shutdown).
The social communication system (ie, social engagement system; see below) involves the myelinated vagus, which serves to foster calm behavioral states by inhibiting sympathetic influences to the heart and dampening the hypothalamic-pituitary-adrenal (HPA) axis.16 The mobilization system is dependent on the functioning of the sympathetic nervous system. The most phylogenetically primitive component, the immobilization system, is dependent on the unmyelinated vagus, which is shared with most vertebrates. With increased neural complexity resulting from phylogenetic development, the organism’s behavioral and affective repertoire is enriched. The three circuits can be conceptualized as dynamic, providing adaptive responses to safe, dangerous, and life-threatening events and contexts.
Only mammals have a myelinated vagus. Unlike the unmyelinated vagus, originating in the dorsal motor nucleus of the vagus with pre- and postganglionic muscarinic receptors, the mammalian myelinated vagus originates in the nucleus ambiguus and has preganglionic nicotinic receptors and postganglionic muscarinic receptors. The unmyelinated vagus is shared with other vertebrates, including reptiles, amphibians, teleosts, and elasmobranchs.
We are now investigating the possibility of extracting different features of the heart rate pattern to dynamically monitor the two vagal systems. Preliminary studies in our laboratory support this possibility. In these studies we have blocked the nicotinic preganglionic receptors with hexamethonium and the muscarinic receptors with atropine. The data were collected from the prairie vole,17 which has a very high ambient vagal tone. These preliminary data demonstrated that, in several animals, nicotinic blockade selectively removes respiratory sinus arrhythmia without dampening the amplitude of the lower frequencies in heart rate variability. In contrast, blocking the muscarinic receptors with atropine removes both the low and respiratory frequencies.
CONSISTENCY WITH JACKSONIAN DISSOLUTION
The three circuits are organized and respond to challenges in a phylogenetically determined hierarchy consistent with the Jacksonian principle of dissolution. Jackson proposed that in the brain, higher (ie, phylogenetically newer) neural circuits inhibit lower (ie, phylogenetically older) neural circuits and “when the higher are suddenly rendered functionless, the lower rise in activity.” 18 Although Jackson proposed dissolution to explain changes in brain function due to damage and illness, the polyvagal theory proposes a similar phylogenetically ordered hierarchical model to describe the sequence of autonomic response strategies to challenges.
The human nervous system, similar to that of other mammals, evolved not solely to survive in safe environments but also to promote survival in dangerous and life-threatening contexts. To accomplish this adaptive flexibility, the human nervous system retained two more primitive neural circuits to regulate defensive strategies (ie, fight–flight and death-feigning behaviors). It is important to note that social behavior, social communication, and visceral homeostasis are incompatible with the neurophysiological states and behaviors promoted by the two neural circuits that support defense strategies. Thus, via evolution, the human nervous system retains three neural circuits, which are in a phylogenetically organized hierarchy. In this hierarchy of adaptive responses, the newest circuit is used first; if that circuit fails to provide safety, the older circuits are recruited sequentially.
Investigation of the phylogeny of regulation of the vertebrate heart11,12,19,20 has led to extraction of four principles that provide a basis for testing of hypotheses relating specific neural mechanisms to social engagement, fight–flight, and death-feigning behaviors:
- There is a phylogenetic shift in the regulation of the heart from endocrine communication to unmyelinated nerves and finally to myelinated nerves.
- There is a development of opposing neural mechanisms of excitation and inhibition to provide rapid regulation of graded metabolic output.
- A face–heart connection evolved as source nuclei of vagal pathways shifted ventrally from the older dorsal motor nucleus to the nucleus ambiguus. This resulted in an anatomical and neurophysiological linkage between neural regulation of the heart via the myelinated vagus and the special visceral efferent pathways that regulate the striated muscles of the face and head, forming an integrated social engagement system (Figure 1; for more details, see Porges7,15).
- With increased cortical development, the cortex exhibits greater control over the brainstem via direct (eg, corticobulbar) and indirect (eg, corticoreticular) neural pathways originating in motor cortex and terminating in the source nuclei of the myelinated motor nerves emerging from the brainstem (eg, specific neural pathways embedded within cranial nerves V, VII, IX, X, and XI), controlling visceromotor structures (ie, heart, bronchi) as well as somatomotor structures (muscles of the face and head).
NEUROCEPTION: CONTEXTUAL CUEING OF ADAPTIVE, MALADAPTIVE PHYSIOLOGICAL STATES
To effectively switch from defensive to social engagement strategies, the mammalian nervous system needs to perform two important adaptive tasks: (1) assess risk, and (2) if the environment is perceived as safe, inhibit the more primitive limbic structures that control fight, flight, or freeze behaviors.
Any stimulus that has the potential for increasing an organism’s experience of safety has the potential of recruiting the evolutionarily more advanced neural circuits that support the prosocial behaviors of the social engagement system.
The nervous system, through the processing of sensory information from the environment and from the viscera, continuously evaluates risk. Since the neural evaluation of risk does not require conscious awareness and may involve subcortical limbic structures,21 the term neuroception22 was introduced to emphasize a neural process, distinct from perception, that is capable of distinguishing environmental (and visceral) features that are safe, dangerous, or life-threatening. In safe environments, autonomic state is adaptively regulated to dampen sympathetic activation and to protect the oxygen-dependent central nervous system, especially the cortex, from the metabolically conservative reactions of the dorsal vagal complex. However, how does the nervous system know when the environment is safe, dangerous, or life-threatening, and which neural mechanisms evaluate this risk?
Environmental components of neuroception
Neuroception represents a neural process that enables humans and other mammals to engage in social behaviors by distinguishing safe from dangerous contexts. Neuroception is proposed as a plausible mechanism mediating both the expression and the disruption of positive social behavior, emotion regulation, and visceral homeostasis.7,22 Neuroception might be triggered by feature detectors involving areas of temporal cortex that communicate with the central nucleus of the amygdala and the periaqueductal gray, since limbic reactivity is modulated by temporal cortex responses to the intention of voices, faces, and hand movements. Thus, the neuroception of familiar individuals and individuals with appropriately prosodic voices and warm, expressive faces translates into a social interaction promoting a sense of safety.
In most individuals (ie, those without a psychiatric disorder or neuropathology), the nervous system evaluates risk and matches neurophysiological state with the actual risk of the environment. When the environment is appraised as being safe, the defensive limbic structures are inhibited, enabling social engagement and calm visceral states to emerge. In contrast, some individuals experience a mismatch and the nervous system appraises the environment as being dangerous even when it is safe. This mismatch results in physiological states that support fight, flight, or freeze behaviors, but not social engagement behaviors. According to the theory, social communication can be expressed efficiently through the social engagement system only when these defensive circuits are inhibited.
Other contributors to neuroception
The features of risk in the environment do not solely drive neuroception. Afferent feedback from the viscera provides a major mediator of the accessibility of prosocial circuits associated with social engagement behaviors. For example, the polyvagal theory predicts that states of mobilization would compromise our ability to detect positive social cues. Functionally, visceral states color our perception of objects and others. Thus, the same features of one person engaging another may result in a range of outcomes, depending on the physiological state of the target individual. If the person being engaged is in a state in which the social engagement system is easily accessible, the reciprocal prosocial interactions are likely to occur. However, if the individual is in a state of mobilization, the same engaging response might be responded to with the asocial features of withdrawal or aggression. In such a state, it might be very difficult to dampen the mobilization circuit and enable the social engagement system to come back on line.
The insula may be involved in the mediation of neuroception, since it has been proposed as a brain structure involved in conveying the diffuse feedback from the viscera into cognitive awareness. Functional imaging experiments have demonstrated that the insula plays an important role in the experience of pain and the experience of several emotions, including anger, fear, disgust, happiness, and sadness. Critchley proposes that internal body states are represented in the insula and contribute to states of subjective feeling, and he has demonstrated that activity in the insula correlates with interoceptive accuracy.23
SUMMARY
The polyvagal theory proposes that the evolution of the mammalian autonomic nervous system provides the neurophysiological substrates for adaptive behavioral strategies. It further proposes that physiological state limits the range of behavior and psychological experience. The theory links the evolution of the autonomic nervous system to affective experience, emotional expression, facial gestures, vocal communication, and contingent social behavior. In this way, the theory provides a plausible explanation for the reported covariation between atypical autonomic regulation (eg, reduced vagal and increased sympathetic influences to the heart) and psychiatric and behavioral disorders that involve difficulties in regulating appropriate social, emotional, and communication behaviors.
The polyvagal theory provides several insights into the adaptive nature of physiological state. First, the theory emphasizes that physiological states support different classes of behavior. For example, a physiological state characterized by a vagal withdrawal would support the mobilization behaviors of fight and flight. In contrast, a physiological state characterized by increased vagal influence on the heart (via myelinated vagal pathways originating in the nucleus ambiguus) would support spontaneous social engagement behaviors. Second, the theory emphasizes the formation of an integrated social engagement system through functional and structural links between neural control of the striated muscles of the face and the smooth muscles of the viscera. Third, the polyvagal theory proposes a mechanism—neuroception— to trigger or to inhibit defense strategies.
- Darwin C. The Expression of Emotions in Man and Animals. New York, NY: D Appleton;1872.
- Langley JN. The Autonomic Nervous System. Cambridge, England: Heffer & Sons; 1921.
- Hess WR. Diencephalon, Autonomic and Extrapyramidal Functions. New York, NY: Grune & Stratton; 1954.
- Porges SW. Vagal tone: a physiologic marker of stress vulnerability. Pediatrics 1992; 90:498–504.
- Hering HE. A functional test of heart vagi in man. Menschen Munchen Medizinische Wochenschrift 1910; 57:1931–1933.
- Porges SW. Respiratory sinus arrhythmia: physiological basis, quantitative methods, and clinical implications. In:Grossman P, Janssen K, Vaitl D, eds. Cardiorespiratory and Cardiosomatic Psychophysiology. New York, NY: Plenum; 1986:101–115.
- Porges SW. The polyvagal perspective. Biol Psychol 2007; 74:116–143.
- Reed SF, Ohel G, David R, Porges SW. A neural explanation of fetal heart rate patterns: a test of the polyvagal theory. Dev Psychobiol 1999; 35:108–118.
- Jordan D, Khalid ME, Schneiderman N, Spyer KM. The location and properties of preganglionic vagal cardiomotor neurones in the rabbit. Pflugers Arch 1982; 395:244–250.
- Katona PG, Jih F. Respiratory sinus arrhythmia: noninvasive measure of parasympathetic cardiac control. J Appl Physiol 1975; 39:801–805.
- Porges SW. Orienting in a defensive world: mammalian modifications of our evolutionary heritage—a polyvagal theory. Psychophysiology 1995; 32:301–318.
- Porges SW. Emotion: An evolutionary by-product of the neural regulation of the autonomic nervous system. Ann N Y Acad Sci 1997; 807:62–77.
- Porges SW. Love: An emergent property of the mammalian autonomic nervous system. Psychoneuroendocrinology 1998; 23:837–861.
- Porges SW. The polyvagal theory: phylogenetic substrates of a social nervous system. Int J Psychophysiol 2001; 42:123–146.
- Porges SW. Social engagement and attachment: a phylogenetic perspective. Ann N Y Acad Sci 2003; 1008:31–47.
- Bueno L, Gue M, Fargeas MJ, Alvinerie M, Junien JL, Fioramonti J. Vagally mediated inhibition of acoustic stress-induced cortisol release by orally administered kappa-opioid substances in dogs. Endocrinology 1989; 124:1788–1793.
- Grippo AJ, Lamb DG, Carter CS, Porges SW. Cardiac regulation in the socially monogamous prairie vole. Physiol Behav 2007; 90:386–393.
- Jackson JH. Evolution and dissolution of the nervous system. In:Taylor J, ed. Selected Writings of John Hughlings Jackson. London: Stapes Press; 1958:45–118.
- Morris JL, Nilsson S. The circulatory system. In:Nilsson S, Holmgren S, eds. Comparative Physiology and Evolution of the Autonomic Nervous System. Chur, Switzerland: Harwood Academic Publishers; 1994.
- Taylor EW, Jordan D, Coote JH. Central control of the cardiovascular and respiratory systems and their interactions in vertebrates. Physiol Rev 1999; 79:855–916.
- Morris JS, Ohman A, Dolan RJ. A subcortical pathway to the right amygdala mediating “unseen” fear. Proc Natl Acad Sci U S A 1999; 96:1680–1685.
- Porges SW. Neuroception: a subconscious system for detecting threat and safety. Zero to Three: Bulletin of the National Center for Clinical Infant Programs 2004; 24( 5):19–24.
- Critchley HD. Neural mechanisms of autonomic, affective, and cognitive integration. J Comp Neurol 2005; 493:154–166.
- Darwin C. The Expression of Emotions in Man and Animals. New York, NY: D Appleton;1872.
- Langley JN. The Autonomic Nervous System. Cambridge, England: Heffer & Sons; 1921.
- Hess WR. Diencephalon, Autonomic and Extrapyramidal Functions. New York, NY: Grune & Stratton; 1954.
- Porges SW. Vagal tone: a physiologic marker of stress vulnerability. Pediatrics 1992; 90:498–504.
- Hering HE. A functional test of heart vagi in man. Menschen Munchen Medizinische Wochenschrift 1910; 57:1931–1933.
- Porges SW. Respiratory sinus arrhythmia: physiological basis, quantitative methods, and clinical implications. In:Grossman P, Janssen K, Vaitl D, eds. Cardiorespiratory and Cardiosomatic Psychophysiology. New York, NY: Plenum; 1986:101–115.
- Porges SW. The polyvagal perspective. Biol Psychol 2007; 74:116–143.
- Reed SF, Ohel G, David R, Porges SW. A neural explanation of fetal heart rate patterns: a test of the polyvagal theory. Dev Psychobiol 1999; 35:108–118.
- Jordan D, Khalid ME, Schneiderman N, Spyer KM. The location and properties of preganglionic vagal cardiomotor neurones in the rabbit. Pflugers Arch 1982; 395:244–250.
- Katona PG, Jih F. Respiratory sinus arrhythmia: noninvasive measure of parasympathetic cardiac control. J Appl Physiol 1975; 39:801–805.
- Porges SW. Orienting in a defensive world: mammalian modifications of our evolutionary heritage—a polyvagal theory. Psychophysiology 1995; 32:301–318.
- Porges SW. Emotion: An evolutionary by-product of the neural regulation of the autonomic nervous system. Ann N Y Acad Sci 1997; 807:62–77.
- Porges SW. Love: An emergent property of the mammalian autonomic nervous system. Psychoneuroendocrinology 1998; 23:837–861.
- Porges SW. The polyvagal theory: phylogenetic substrates of a social nervous system. Int J Psychophysiol 2001; 42:123–146.
- Porges SW. Social engagement and attachment: a phylogenetic perspective. Ann N Y Acad Sci 2003; 1008:31–47.
- Bueno L, Gue M, Fargeas MJ, Alvinerie M, Junien JL, Fioramonti J. Vagally mediated inhibition of acoustic stress-induced cortisol release by orally administered kappa-opioid substances in dogs. Endocrinology 1989; 124:1788–1793.
- Grippo AJ, Lamb DG, Carter CS, Porges SW. Cardiac regulation in the socially monogamous prairie vole. Physiol Behav 2007; 90:386–393.
- Jackson JH. Evolution and dissolution of the nervous system. In:Taylor J, ed. Selected Writings of John Hughlings Jackson. London: Stapes Press; 1958:45–118.
- Morris JL, Nilsson S. The circulatory system. In:Nilsson S, Holmgren S, eds. Comparative Physiology and Evolution of the Autonomic Nervous System. Chur, Switzerland: Harwood Academic Publishers; 1994.
- Taylor EW, Jordan D, Coote JH. Central control of the cardiovascular and respiratory systems and their interactions in vertebrates. Physiol Rev 1999; 79:855–916.
- Morris JS, Ohman A, Dolan RJ. A subcortical pathway to the right amygdala mediating “unseen” fear. Proc Natl Acad Sci U S A 1999; 96:1680–1685.
- Porges SW. Neuroception: a subconscious system for detecting threat and safety. Zero to Three: Bulletin of the National Center for Clinical Infant Programs 2004; 24( 5):19–24.
- Critchley HD. Neural mechanisms of autonomic, affective, and cognitive integration. J Comp Neurol 2005; 493:154–166.